Introduction
The sensations of touch and pain get perceived by cutaneous low threshold mechanoreceptors (LTMs) in the skin. These cells convert tactile stimuli into membrane action potentials that then trigger neuronal action potentials. These LTMs can adapt and transmit sensations to sensory neurons, which project to the spinal cord at the dorsal horn to relay light touch or to the lateral spinothalamic tract to convey pain.[1][2] For somatosensory perception, the LTMs lie within the dorsal root ganglia. The dorsal root ganglion neurons are pseudo-unipolar; one axonal branch goes to the periphery and the other synapses in the spinal cord. LTMs can have slowly adapting receptors for sustained indentation and rapidly adapting receptors that respond to the start and endpoint of indentation.[3]
Structure and Function
Sensory neurons are classified based on axon diameter, cell body size, axonal conduction velocities, and degree of myelination.[2] C fibers are the smallest, unmyelinated, have slow conduction velocities (0.2 to 2 m/s), and are the most abundant. These fibers typically transmit sustained slow sensations. A-delta fibers have a medium-sized cell body, are lightly myelinated, and have a moderate conduction velocity (5 to 30 m/s). They are responsible for quick sensations and prickling pain. A-beta fibers have a large cell body, are heavily myelinated, and have rapid conduction velocities (16 to 100 m/s). A-beta fibers are light-touch receptors that have low mechanical thresholds while A-delta and C fibers are nociceptors that respond to noxious mechanical, hot, or cold stimuli. These different fibers comprise different touch end organs that respond to various tactile stimuli.[4]
There are two major types of mammalian skin; glabrous (non-hairy) and hairy skin. Glabrous skin is specific for discriminative touch and is present in the hands and feet. This skin gives proper grip control and can determine texture and shape. Hairy skin covers the other 90% of our body and is closely related to both pleasurable sensations, like stroking, and pain, such as hair pulling.[3] Four mechanosensory end organs are found in glabrous skin: Ruffini endings, Meissner corpuscles, Pacinian corpuscles, and Merkel discs. Both Meissner corpuscles and Pacinian corpuscles are rapidly adapting while Merkel cells and Ruffini endings are slowly adapting nerve fibers.[1]
Embryology
The mechanosensory neurons are periphery neurons that stretch to the spinal cord to connect to the central nervous system. All of the low threshold mechanoreceptors come from precursor neural crest cells in early development. These transient, multipotent cells move from the dorsal neural tube to the periphery in the dorsal root ganglia (DRG) in parallel with skin development. The skin originates from the ectoderm, and the mesoderm, and these axons from the DRG grow to meet the skin. Neurons that specialize early express Ngn2; these include A-beta and A-delta afferents. Later development expresses Ngn1, creating C afferents.[5]
Blood Supply and Lymphatics
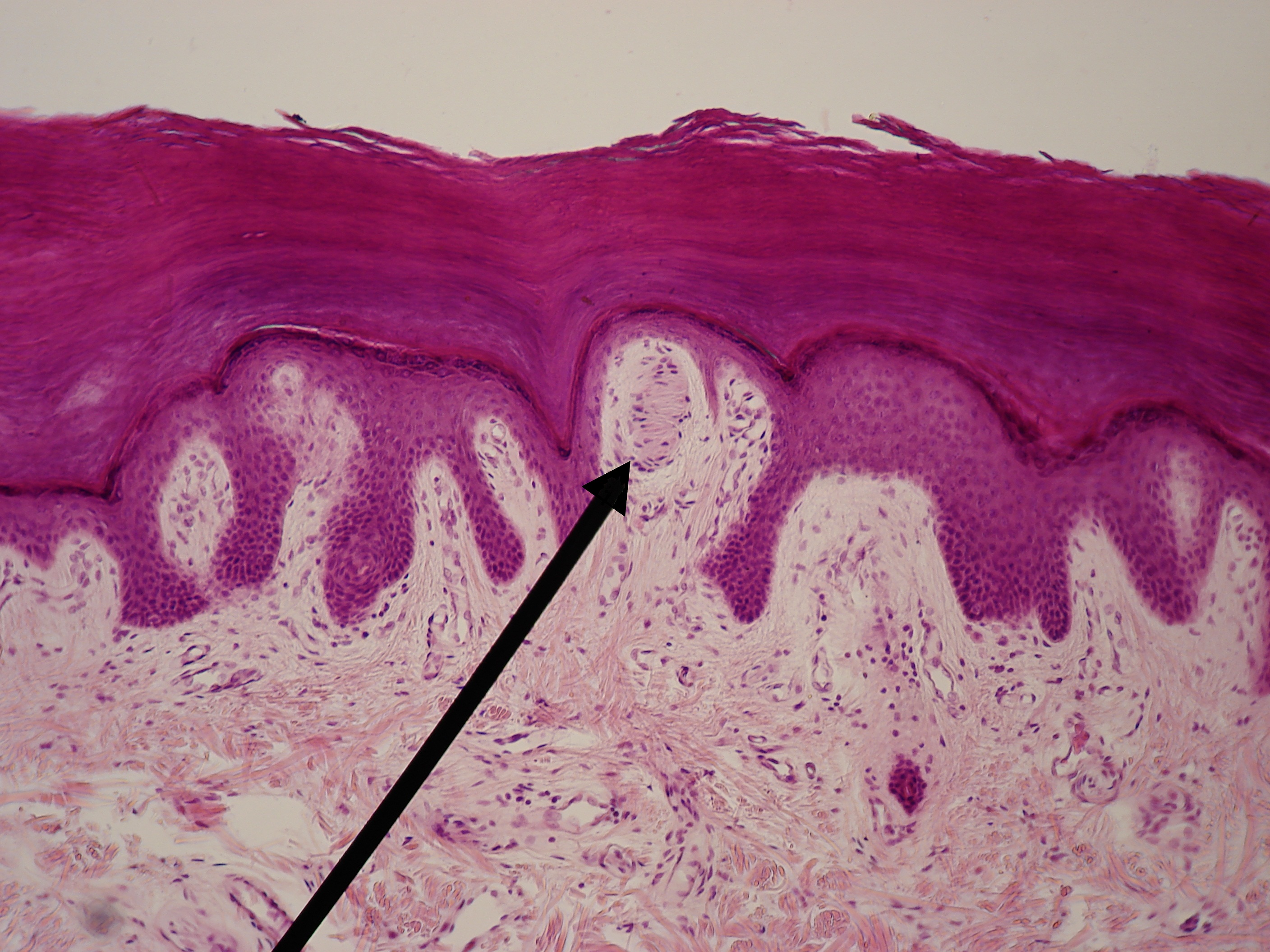
The epidermal layer of the skin does not have blood or lymphatic vessels. The dermis has a superficial and deep horizontal vascular plexus. Capillaries arise from the superficial plexus and terminate close to the epidermal layer. Lymphatics also make two plexuses near the blood vessels; the superficial lymphatic plexus moves through the dermal papillae and descends into the lower dermis. The lymphatic vessels do not come as close to the epidermis as the blood vessels. These vessels provide nutrients, discard waste, and produce inflammation for the nerves found in the skin.
Nerves
Merkel cells appear in the basal layer of the epidermis and use A-beta nerves to detect indentation and tactile stimuli to reconstruct spatial images. Merkel cells are involved in Merkel cell-neurite complexes located in skin areas with high tactile acuity, such as vibrissal follicles, glabrous fingerpads, and highly sensitive areas of hairy skin called touch domes. Merkel cells use slowly adapting type I (SAI-LTMs) responses to represent sustained pressure.[6] These signals can accurately interpret and recreate object features. Additionally, there are fast; touch-evoked mechanotransduction currents in the Merkel cell-neurite complex that send high-frequency signals for acute stimuli like grating movements.[7] Merkel cell-neurite complexes send currents through calcium-permeable cation channels (Piezo2) that can be activated by skin swelling and membrane stretch, leading to the release of norepinephrine, which binds to B-adrenergic receptors on the sensory neuron.[8] These cells exhibit high sensitivity for spatial resolution, therefore perfect for determining stimulus position and velocity.[6]
Ruffini endings are deep in the dermis and are postulated to have slow adapting type II mechanoreceptors (SAII-LTMs), but researchers have not fully studied them. They are large, thin spindle-shaped cylinders comprised of perineural tissue, such as collagen fibers and Swann cells, and have a fluid-filled capsule space as the inner core. They have low-threshold regions and sense skin stretch by interacting with collagen fibers.[9] Ruffini corpuscles do not possess specialized transducer cells, leading many to believe that the neuron itself acts as the mechano-electric transducer.[10]
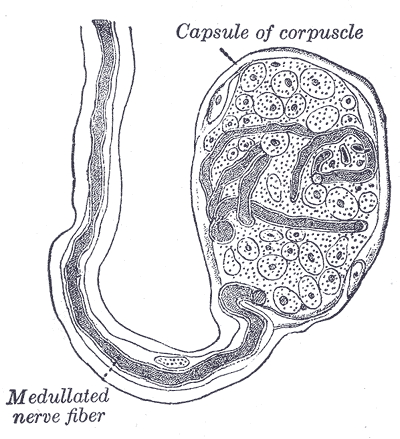
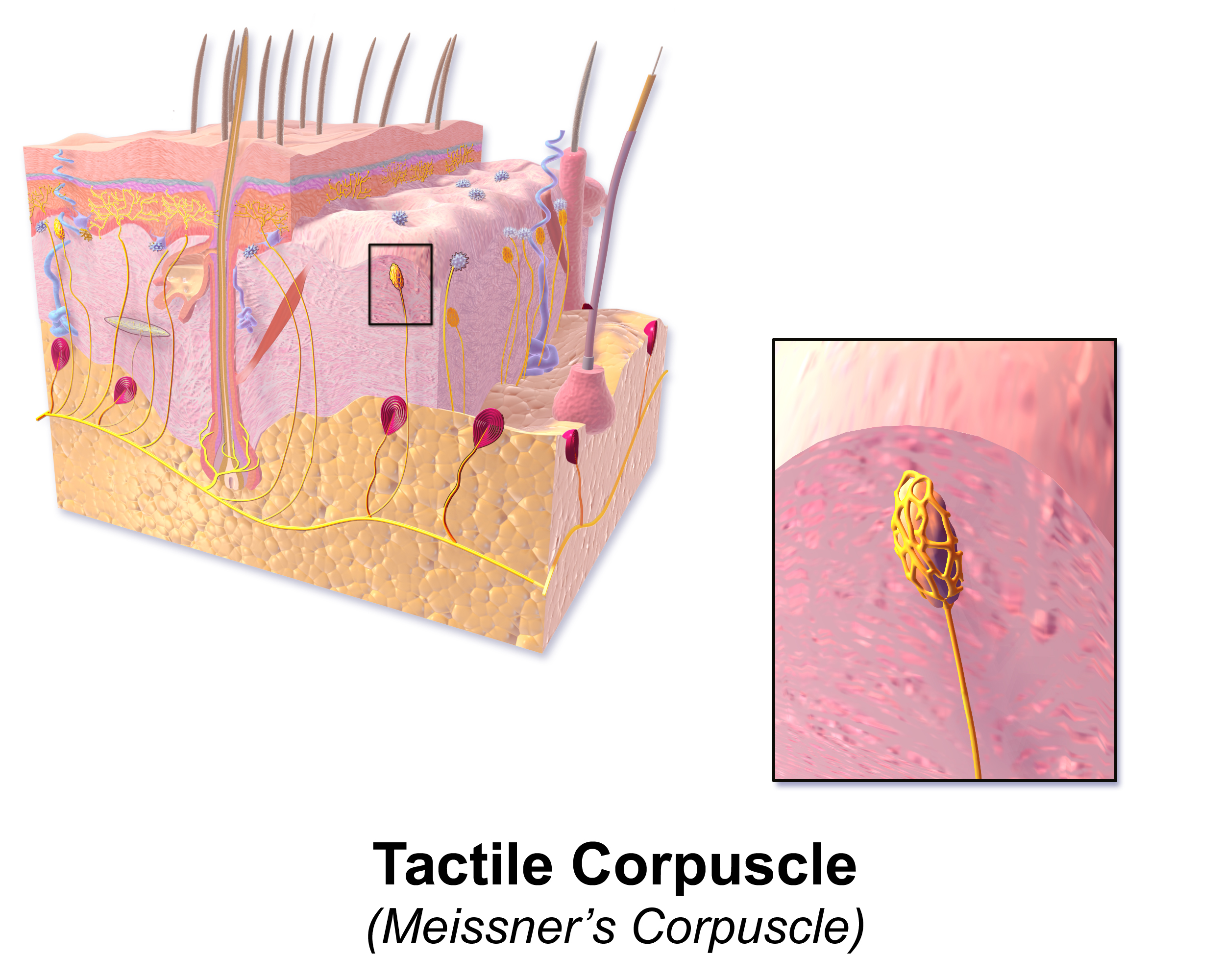
Meissner corpuscles use A-beta as well to detect skin movement through type I rapidly adapting mechanoreceptors (RAI-LTMs). These are tuned to detect rapid movement, or flutter, and to control grip. They are in the dermal papillae layer of the skin and organize into flattened, horizontal lamellar cells enmeshed in connective tissue. These receptors have four times the sensitivity as Merkel cells but lack the ability to determine spatial differences. Their location is at the fingertips and soles of the feet. When feeling tactile indentation, the collagen fibers around the Meissner corpuscles stretch and cause the corpuscle to deform, triggering multiple action potentials. When the pressure is released, the corpuscle regains its shape, and again volleys off multiple action potentials. RAI-LTMs and SAI-LTMs most likely combine to form a complete picture of tactile stimulation.[11]
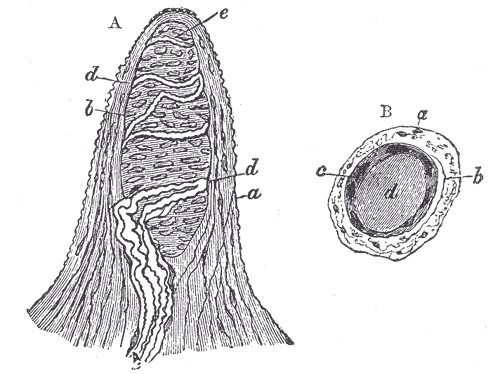
Pacinian corpuscles detect high-frequency vibration and are in the dermis of glabrous skin. These cells use type II, rapidly adapting mechanoreceptors (RAII-LTMs). The corpuscles are large and oval-shaped with onion-shaped lamellar cells surrounding an A-beta nerve ending. RAII-LTMs are the most sensitive and cover large areas of skin. This arrangement makes them very adept at picking up vibrations with great accuracy but unable to localize the sensation with any spatial acuity. These nerve organs allow us to interpret the change in vibration as our hands manipulate objects.[12]
The pilosebaceous unit, containing hair follicles, works to produce hair fibers and oils, participates in wound healing, and holds reserves of immune cells and pigment-producing cells. The isthmus and upper bulge region of a hair follicle contains a piloneural collar. The piloneural collar contains mechanoreceptors that transmit sensation through the hair follicle. Different hair follicle types are defined by their length, hair shaft thickness, and kinks. The types are called zigzag, auchene/awl, and guard.[13][14] Piloneural collars can contain all three types of sensory neurons; A-beta, A-delta, or C fibers. A-beta fibers were previously discussed in conjunction with Merkel cells touch domes. A-delta cells can be hair follicle specific, sometimes called D-Hair units, and are the most sensitive hair follicle mechanoreceptor. Some A-delta fibers are circumferentially wrapped around the base of hair follicles, while others have longitudinal lanceolate endings. These D-Hair units are associated with the awl/auchene and zigzag hair follicles and detect the movements of the small sinuses and down hairs in mammals. It is still unknown how these cells affect touch perception. C fibers are associated with painful stimuli, pleasurable touches, and sensations such as tickling. C fibers are the most numerous and are activated by indentation that moves slowly across their field, called caressing touches. These fibers are only found in hairy skin and are postulated to play a role in emotional touch.[3][9]
Clinical Significance
Lesions at multiple levels can affect the chain of neurons and synapses that transmit the sensations of touch from the skin to the central nervous system. The peripheral nerves can get severed due to trauma or demyelinated in several autoimmune diseases; these nerves can regrow as long as the cell body of the nerve is intact. These nerves can regrow at 2 mm/day in small nerves or 5 mm/day in large nerves. As previously discussed, the dorsal horn relays sensation to light touch and vibration while the lateral spinothalamic tract relays pain sensations. If a spinal cord lesion occurs in either of these tracts, the spinal nerve levels below the lesions will lose sensation.