
Introduction
Although the DNA helical diameter is only 2 nm, the entire DNA strand in a single cell will stretch roughly 2 meters when completely unwound.[1] The entire DNA strand must fit within the nucleus of a cell, so it must be very tightly packaged to fit. This is accomplished by wrapping the DNA around structural histone proteins, which act as scaffolding for the DNA to be coiled around. The entire structure is called a nucleosome, each of which includes an octamer of histone proteins and 146 to 147 base pairs of DNA.[2][3] The millions of nucleosomes tightly coil the continuous DNA strand into chromatin which is further condensed into the chromosome classically visualized during cell division.
The nucleosome structure was first described in 1974 by Roger Kornberg who, with evidence from biochemical experiments, X-ray diffraction studies, and electron microscopy images, proposed the nucleosome was comprised of repeating units of eight histone proteins and about 200 DNA base pairs.[4] In the same year, Markus Noll gave a visually interpretable result to understand how DNA wraps around the nucleosomes.[5] The results of his experiments showed that DNA wrapped was cleaved when exposed to DNAse I, which suggested that the enzyme must have had access to the DNA. The results of this experiment favored the theory that DNA was wound on the outside of the nucleosome unit and supported the nucleosome structure proposed by Kornberg, with each nucleosome consisting of approximately 200 DNA base pairs.
The tight structure of chromatin brings about the problem of accessibility to the DNA by enzymes involved in DNA replication and transcription. Chromatin exists in one of two states: heterochromatin, which is condensed and allows little access by transcription enzymes, and euchromatin, which is loose to allow for interaction with transcription enzymes. The transition between these two states is determined by interactions between the DNA and histone proteins via post-translational modifications to the histone proteins like methylation and acetylation.[6] Methylation generally increases interactions between the DNA and histone, thus suppressing gene expression, whereas acetylation will loosen interactions resulting in greater access by transcription enzymes resulting in increased gene expression.[7] The post-translational modifications to histone proteins underlie the mechanisms of epigenetics, which are defined as alterations to gene expression without changes to the DNA sequence.[8][9]
The ability for DNA packaging to be modified at various stages of the cell cycle is important in both DNA replication and cell division as well as transcription. Replication occurs at many origins of replication throughout the DNA strand to accelerate the replication of the entire genome, with each origin separated by approximately 100,000 base pairs.[10] The DNA does not interact with histones during this process to allow for the propagation of the polymerase enzymes. However, when the process is complete, the DNA must reintegrate with the histones to reform nucleosomes and eventually the supercoiled chromosome structure during mitosis. Following cell division, the DNA must again separate from the histone proteins to undergo transcription. This capability for the DNA-histone interactions to be modulated is crucial for the proper growth and function of a cell with malfunctions contributing to disease like hypermethylation in cancer.[11]
Molecular Level
Register For Free And Read The Full Article
 Search engine and full access to all medical articles
Search engine and full access to all medical articles- 10 free questions in your specialty
- Free CME/CE Activities
- Free daily question in your email
- Save favorite articles to your dashboard
- Emails offering discounts
Learn more about a Subscription to StatPearls Point-of-Care
Molecular Level
All DNA is packaged as structures called nucleosomes which are the basic unit of chromatin.[3] Nucleosomes are composed of double-stranded DNA wrapped around an octamer of histone proteins, including two of each of the following: H2A, H2B, H3, and H4. The octamer structure is a four-helix bundle comprised of two H3-H4 and two H2A-H2B dimers.[12] An additional histone protein, H1, is not part of the core histone octamer but plays a role in nucleosome stability by acting as a linker protein to further stabilize the nucleosome.[13] Histone proteins have high lysine and arginine content which makes them positively charged.[14] DNA is largely negatively charged, thus favoring interactions with positively charged histone proteins that stabilize the nucleosome.[15] Histone stabilization occurs via numerous protein-protein interactions, hydrogen bonding, and electrostatic forces.[2][16][17]
Further DNA binding proteins, known as non-histone proteins, are a large group of heterogeneous proteins that play a role in the organization and compaction of the chromosome into higher-order structures. The H1 protein is essential in these higher-order structures.[18][19] Secondary structures to chromatin are demonstrated by the solenoid model and the zigzag model.[20] The solenoid model consists of tightly wound nucleosomes in a regular, spiral configuration containing 6 nucleosomes per turn.[21] The zigzag model is a bit looser form of chromatin with irregular configuration. In this model, nucleosomes have little face-to-face contact. In both the solenoid and the zigZag model, fibers are 30-nanometer in size.[22]
From the solenoid size, chromatin is further packaged and condensed into chromosomes. These regions can be visualized microscopically through various stains applied to chromosomes during mitosis that are best seen during metaphase.[23] Even though DNA appears to be unorganized intranuclearly during interphase, there is still a significant organization of the structure and partitioning of different chromosomal material within the nucleus. The DNA from individual chromosomes is not intertwined with other chromosomes but remains in specific regions of the nucleus called chromosome territories.[24] These territories can help bring different genes into a spatial relationship with one another, which is felt to be an important regulator of gene expression.
In addition to the need for systematic compaction of DNA for replication and cell division, it is important for the interphase cell to have its DNA organized within the nucleus. This organization helps to section the DNA into different areas of cell expression, among other functions. The nucleus consists of a nuclear double-layered membrane matrix compiled of different types of proteins that ensure nuclear stability and facilitate nuclear organization. This organization is by no means static, and a plethora of complicated mechanisms will change DNA expression temporally and geographically within the body. The nuclear lamina is under the inner membrane of the nucleus, where scaffolding proteins and matrix attachment proteins are present. Eukaryotic DNA is organized into loops, which can vary from 25 to 200 base pairs long. Within the actual genetic code of DNA, specific sequences allow for the attachment of matrix attachment regions (MARs) and scaffold attachment regions (SARs) along the nuclear lamina.[25] SARs are where the DNA is bound to the matrix or scaffold of the chromosome, and the MARs are attached to the nuclear matrix creating these radial loops.[26] These regions have been shown to be involved in gene expression.[27]
Clinical Significance
The clinical significance of aberration in DNA organization is broad-reaching. This is highlighted by many disorders of histone methylation, including alpha thalassemia X-linked intellectual disability syndrome, Rubinstein-Taybi syndrome, Coffin-Lowry syndrome, and Rett Syndrome.[28][29][30][31][32][33]
DNA packaging around histones is directly related to gene expression and is the foundation of the field of epigenetics. The extent to which epigenetics contributes to clinical presentations is still actively being determined, but the present body of evidence suggests that variable gene expression contributes to a range of pathologic processes involved in cancers, neurological disorders, and even consciousness, among many other disease states.[34][35][36] The mechanisms by which epigenetic changes contribute to disease are complex but generally are related to either increased or decreased expression of specific genes. Two common mechanisms include acetylation of the histone protein to increase gene expression and methylation of histone lysine residues to decrease gene expression. Additional post-translational modifications to histone proteins that affect gene expression include ubiquitination, phosphorylation, and ADP-ribosylation.[34] Each of these modifications will either strengthen or disrupt the DNA-histone interactions to allow for subsequent changes in accessibility by transcriptional enzymes.
Important to the field of epigenetics is the fact that the changes associated with disease states are reversible, unlike mutations in the DNA sequence itself. With this in mind, the increased understanding of epigenetics has elucidated the potential to treat certain conditions by preventing or reversing the post-translational modifications to histone proteins. For example, methylation of a tumor suppressor gene or deacetylation of a histone protein associated with an oncogene likely contributes to carcinogenesis, so drugs that inhibit these processes have been trialed as potential cancer treatments.[37] The FDA has already approved multiple drugs for treating myelodysplastic syndrome by inhibiting DNA methyltransferases resulting in a decrease in DNA hypermethylation associated with these cancers.[38] Drugs to reverse post-translational histone modifications to methylation and acetylation status have also been trialed for cancers including melanoma, lung cancer, and colorectal cancer, with results showing some promise though many trials are still ongoing.[39][40]
Media
(Click Image to Enlarge)
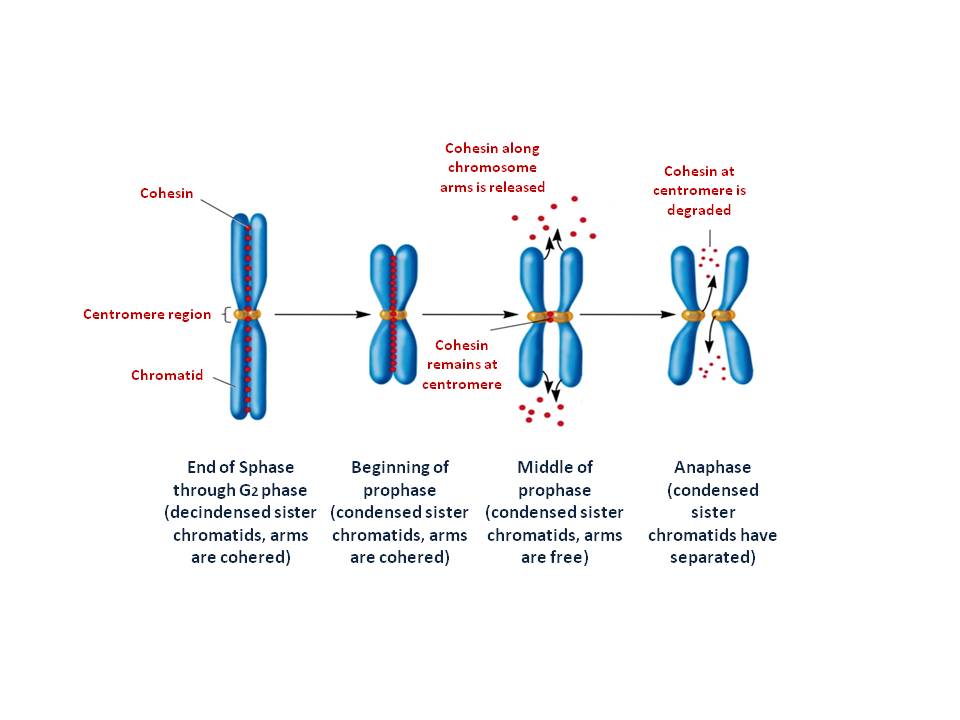
The two multi protein complexes help to form and organize metaphase chromosomes and cohesin which keeps the two sister chromatid alignment
Contributed by Nora Al Aboud, PhD (Genetics)
References
Travers A, Muskhelishvili G. DNA structure and function. The FEBS journal. 2015 Jun:282(12):2279-95. doi: 10.1111/febs.13307. Epub 2015 Jun 2 [PubMed PMID: 25903461]
Luger K, Mäder AW, Richmond RK, Sargent DF, Richmond TJ. Crystal structure of the nucleosome core particle at 2.8 A resolution. Nature. 1997 Sep 18:389(6648):251-60 [PubMed PMID: 9305837]
Mariño-Ramírez L, Kann MG, Shoemaker BA, Landsman D. Histone structure and nucleosome stability. Expert review of proteomics. 2005 Oct:2(5):719-29 [PubMed PMID: 16209651]
Level 3 (low-level) evidenceKornberg RD. Chromatin structure: a repeating unit of histones and DNA. Science (New York, N.Y.). 1974 May 24:184(4139):868-71 [PubMed PMID: 4825889]
Level 3 (low-level) evidenceNoll M. Internal structure of the chromatin subunit. Nucleic acids research. 1974 Nov:1(11):1573-8 [PubMed PMID: 10793712]
Level 3 (low-level) evidenceTessarz P, Kouzarides T. Histone core modifications regulating nucleosome structure and dynamics. Nature reviews. Molecular cell biology. 2014 Nov:15(11):703-8. doi: 10.1038/nrm3890. Epub 2014 Oct 15 [PubMed PMID: 25315270]
Gallinari P, Di Marco S, Jones P, Pallaoro M, Steinkühler C. HDACs, histone deacetylation and gene transcription: from molecular biology to cancer therapeutics. Cell research. 2007 Mar:17(3):195-211 [PubMed PMID: 17325692]
Level 3 (low-level) evidenceKatan-Khaykovich Y, Struhl K. Heterochromatin formation involves changes in histone modifications over multiple cell generations. The EMBO journal. 2005 Jun 15:24(12):2138-49 [PubMed PMID: 15920479]
Pinel C, Prainsack B, McKevitt C. Markers as mediators: A review and synthesis of epigenetics literature. BioSocieties. 2019 May 10:13(1):276-303. doi: 10.1057/s41292-017-0068-x. Epub 2017 Sep 18 [PubMed PMID: 31105763]
Fangman WL, Brewer BJ. A question of time: replication origins of eukaryotic chromosomes. Cell. 1992 Oct 30:71(3):363-6 [PubMed PMID: 1423601]
Level 3 (low-level) evidenceKulis M, Esteller M. DNA methylation and cancer. Advances in genetics. 2010:70():27-56. doi: 10.1016/B978-0-12-380866-0.60002-2. Epub [PubMed PMID: 20920744]
Level 3 (low-level) evidenceMcGinty RK, Tan S. Nucleosome structure and function. Chemical reviews. 2015 Mar 25:115(6):2255-73. doi: 10.1021/cr500373h. Epub 2014 Dec 12 [PubMed PMID: 25495456]
Kalashnikova AA, Rogge RA, Hansen JC. Linker histone H1 and protein-protein interactions. Biochimica et biophysica acta. 2016 Mar:1859(3):455-61. doi: 10.1016/j.bbagrm.2015.10.004. Epub 2015 Oct 8 [PubMed PMID: 26455956]
Smith BC, Denu JM. Chemical mechanisms of histone lysine and arginine modifications. Biochimica et biophysica acta. 2009 Jan:1789(1):45-57. doi: 10.1016/j.bbagrm.2008.06.005. Epub 2008 Jun 14 [PubMed PMID: 18603028]
Level 3 (low-level) evidenceDueva R, Akopyan K, Pederiva C, Trevisan D, Dhanjal S, Lindqvist A, Farnebo M. Neutralization of the Positive Charges on Histone Tails by RNA Promotes an Open Chromatin Structure. Cell chemical biology. 2019 Oct 17:26(10):1436-1449.e5. doi: 10.1016/j.chembiol.2019.08.002. Epub 2019 Aug 22 [PubMed PMID: 31447351]
Davey CA, Sargent DF, Luger K, Maeder AW, Richmond TJ. Solvent mediated interactions in the structure of the nucleosome core particle at 1.9 a resolution. Journal of molecular biology. 2002 Jun 21:319(5):1097-113 [PubMed PMID: 12079350]
Level 3 (low-level) evidenceRohs R, West SM, Sosinsky A, Liu P, Mann RS, Honig B. The role of DNA shape in protein-DNA recognition. Nature. 2009 Oct 29:461(7268):1248-53. doi: 10.1038/nature08473. Epub [PubMed PMID: 19865164]
Level 3 (low-level) evidenceTremethick DJ. Higher-order structures of chromatin: the elusive 30 nm fiber. Cell. 2007 Feb 23:128(4):651-4 [PubMed PMID: 17320503]
Level 3 (low-level) evidenceThoma F, Koller T. Influence of histone H1 on chromatin structure. Cell. 1977 Sep:12(1):101-7 [PubMed PMID: 561660]
Level 3 (low-level) evidenceChen P, Li G. Dynamics of the higher-order structure of chromatin. Protein & cell. 2010 Nov:1(11):967-71. doi: 10.1007/s13238-010-0130-y. Epub [PubMed PMID: 21153512]
Level 3 (low-level) evidenceKruithof M, Chien FT, Routh A, Logie C, Rhodes D, van Noort J. Single-molecule force spectroscopy reveals a highly compliant helical folding for the 30-nm chromatin fiber. Nature structural & molecular biology. 2009 May:16(5):534-40. doi: 10.1038/nsmb.1590. Epub 2009 Apr 19 [PubMed PMID: 19377481]
Level 3 (low-level) evidenceLuger K, Dechassa ML, Tremethick DJ. New insights into nucleosome and chromatin structure: an ordered state or a disordered affair? Nature reviews. Molecular cell biology. 2012 Jun 22:13(7):436-47. doi: 10.1038/nrm3382. Epub 2012 Jun 22 [PubMed PMID: 22722606]
Level 3 (low-level) evidenceKireeva N, Lakonishok M, Kireev I, Hirano T, Belmont AS. Visualization of early chromosome condensation: a hierarchical folding, axial glue model of chromosome structure. The Journal of cell biology. 2004 Sep 13:166(6):775-85 [PubMed PMID: 15353545]
Level 3 (low-level) evidenceCremer T, Cremer M. Chromosome territories. Cold Spring Harbor perspectives in biology. 2010 Mar:2(3):a003889. doi: 10.1101/cshperspect.a003889. Epub [PubMed PMID: 20300217]
Level 3 (low-level) evidenceCockerill PN, Garrard WT. Chromosomal loop anchorage of the kappa immunoglobulin gene occurs next to the enhancer in a region containing topoisomerase II sites. Cell. 1986 Jan 31:44(2):273-82 [PubMed PMID: 3002631]
Level 3 (low-level) evidenceHeng HH, Goetze S, Ye CJ, Liu G, Stevens JB, Bremer SW, Wykes SM, Bode J, Krawetz SA. Chromatin loops are selectively anchored using scaffold/matrix-attachment regions. Journal of cell science. 2004 Mar 1:117(Pt 7):999-1008 [PubMed PMID: 14996931]
Level 3 (low-level) evidenceNarwade N, Patel S, Alam A, Chattopadhyay S, Mittal S, Kulkarni A. Mapping of scaffold/matrix attachment regions in human genome: a data mining exercise. Nucleic acids research. 2019 Aug 22:47(14):7247-7261. doi: 10.1093/nar/gkz562. Epub [PubMed PMID: 31265077]
Bouazzi H, Thakur S, Trujillo C, Alwasiyah MK, Munnich A. Novel ATRX gene damaging missense mutation c.6740A}C segregates with profound to severe intellectual deficiency without alpha thalassaemia. The Indian journal of medical research. 2016 Jan:143(1):43-8. doi: 10.4103/0971-5916.178589. Epub [PubMed PMID: 26997013]
Iwase S, Bérubé NG, Zhou Z, Kasri NN, Battaglioli E, Scandaglia M, Barco A. Epigenetic Etiology of Intellectual Disability. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2017 Nov 8:37(45):10773-10782. doi: 10.1523/JNEUROSCI.1840-17.2017. Epub [PubMed PMID: 29118205]
Berdasco M, Esteller M. Genetic syndromes caused by mutations in epigenetic genes. Human genetics. 2013 Apr:132(4):359-83. doi: 10.1007/s00439-013-1271-x. Epub 2013 Jan 31 [PubMed PMID: 23370504]
Level 3 (low-level) evidenceWada T. [X-linked alpha-thalassemia/mental retardation syndrome]. Rinsho byori. The Japanese journal of clinical pathology. 2009 Apr:57(4):382-90 [PubMed PMID: 19489441]
Level 3 (low-level) evidenceOlson CO, Pejhan S, Kroft D, Sheikholeslami K, Fuss D, Buist M, Ali Sher A, Del Bigio MR, Sztainberg Y, Siu VM, Ang LC, Sabourin-Felix M, Moss T, Rastegar M. MECP2 Mutation Interrupts Nucleolin-mTOR-P70S6K Signaling in Rett Syndrome Patients. Frontiers in genetics. 2018:9():635. doi: 10.3389/fgene.2018.00635. Epub 2018 Dec 19 [PubMed PMID: 30619462]
Urdinguio RG, Sanchez-Mut JV, Esteller M. Epigenetic mechanisms in neurological diseases: genes, syndromes, and therapies. The Lancet. Neurology. 2009 Nov:8(11):1056-72. doi: 10.1016/S1474-4422(09)70262-5. Epub [PubMed PMID: 19833297]
Level 3 (low-level) evidenceRozek LS, Dolinoy DC, Sartor MA, Omenn GS. Epigenetics: relevance and implications for public health. Annual review of public health. 2014:35():105-22. doi: 10.1146/annurev-publhealth-032013-182513. Epub [PubMed PMID: 24641556]
Level 3 (low-level) evidenceSen N. Epigenetic regulation of memory by acetylation and methylation of chromatin: implications in neurological disorders, aging, and addiction. Neuromolecular medicine. 2015 Jun:17(2):97-110. doi: 10.1007/s12017-014-8306-x. Epub 2014 Apr 29 [PubMed PMID: 24777294]
Level 3 (low-level) evidenceHudson AJ. Consciousness and cell memory: a dynamic epigenetic interrelationship. The Canadian journal of neurological sciences. Le journal canadien des sciences neurologiques. 2011 Sep:38(5):681-8 [PubMed PMID: 21856569]
Perri F, Longo F, Giuliano M, Sabbatino F, Favia G, Ionna F, Addeo R, Della Vittoria Scarpati G, Di Lorenzo G, Pisconti S. Epigenetic control of gene expression: Potential implications for cancer treatment. Critical reviews in oncology/hematology. 2017 Mar:111():166-172. doi: 10.1016/j.critrevonc.2017.01.020. Epub 2017 Feb 4 [PubMed PMID: 28259291]
Derissen EJ, Beijnen JH, Schellens JH. Concise drug review: azacitidine and decitabine. The oncologist. 2013:18(5):619-24. doi: 10.1634/theoncologist.2012-0465. Epub 2013 May 13 [PubMed PMID: 23671007]
Haas NB,Quirt I,Hotte S,McWhirter E,Polintan R,Litwin S,Adams PD,McBryan T,Wang L,Martin LP,vonMehren M,Alpaugh RK,Zweibel J,Oza A, Phase II trial of vorinostat in advanced melanoma. Investigational new drugs. 2014 Jun; [PubMed PMID: 24464266]
Cheng Y, He C, Wang M, Ma X, Mo F, Yang S, Han J, Wei X. Targeting epigenetic regulators for cancer therapy: mechanisms and advances in clinical trials. Signal transduction and targeted therapy. 2019:4():62. doi: 10.1038/s41392-019-0095-0. Epub 2019 Dec 17 [PubMed PMID: 31871779]
Level 3 (low-level) evidence