Introduction
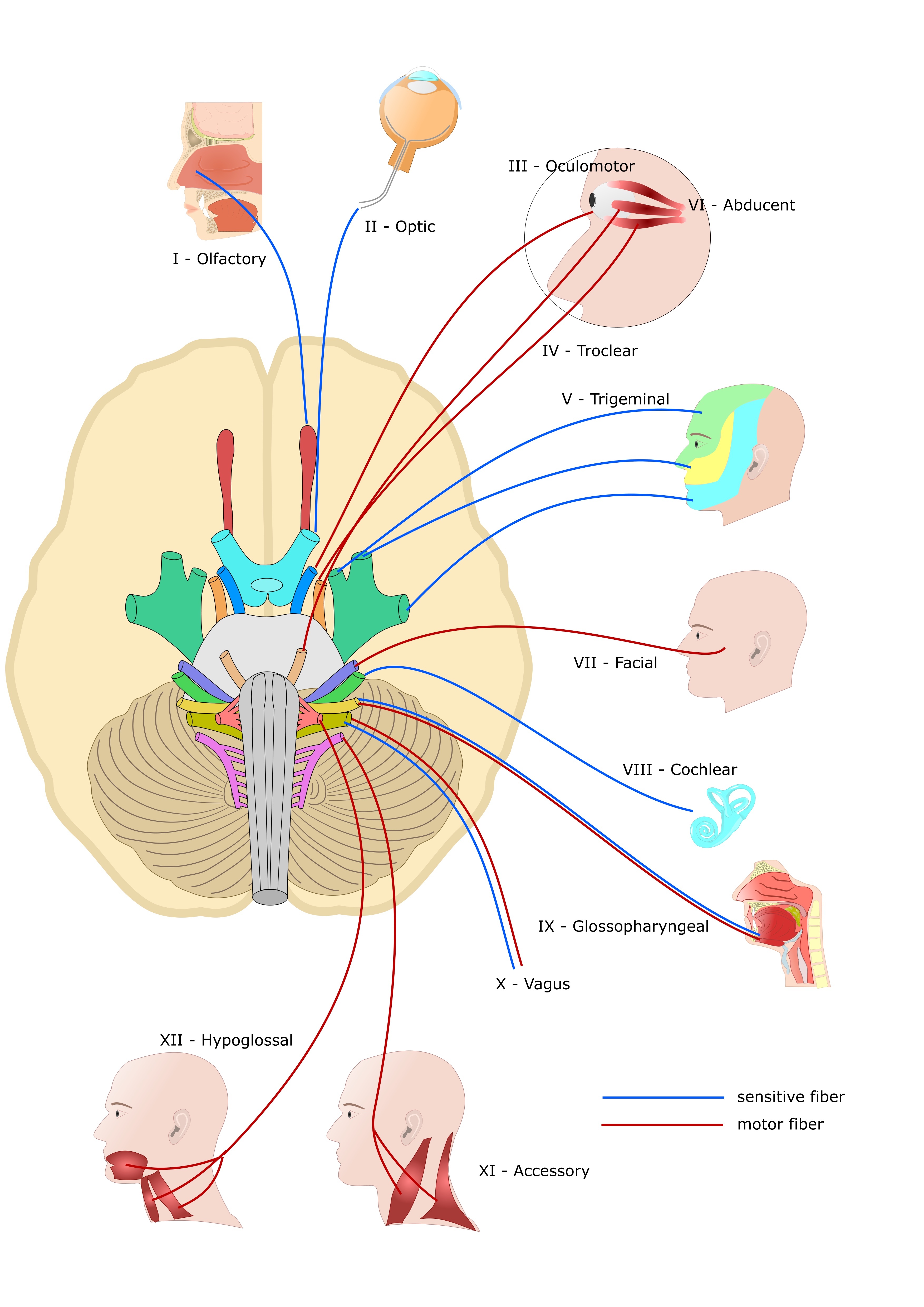
The cranial nerves provide afferent and efferent innervation principally to the structures of the head and neck. Unlike spinal nerves, whose roots are neural fibers from the spinal grey matter, cranial nerves are composed of the neural processes associated with distinct brainstem nuclei and cortical structures. Unlike the spinal nerves, cranial nerve nuclei are functionally organized into distinct nuclei within the brainstem. Typically, the more posterior and lateral nuclei tend to be sensory, and the more anterior tend to be motor. Cranial nerves I (olfactory), II (optic), and VIII (vestibulocochlear) are considered purely afferent. Cranial nerves III (oculomotor), IV (trochlear), VI (abducens), XI (spinal accessory), and XII (hypoglossal) are purely efferent. The remaining cranial nerves, V (trigeminal), VII (facial), IX (glossopharyngeal), and X (vagus), are functionally mixed (sensory and motor).[1] See Image. Cranial Nerves.
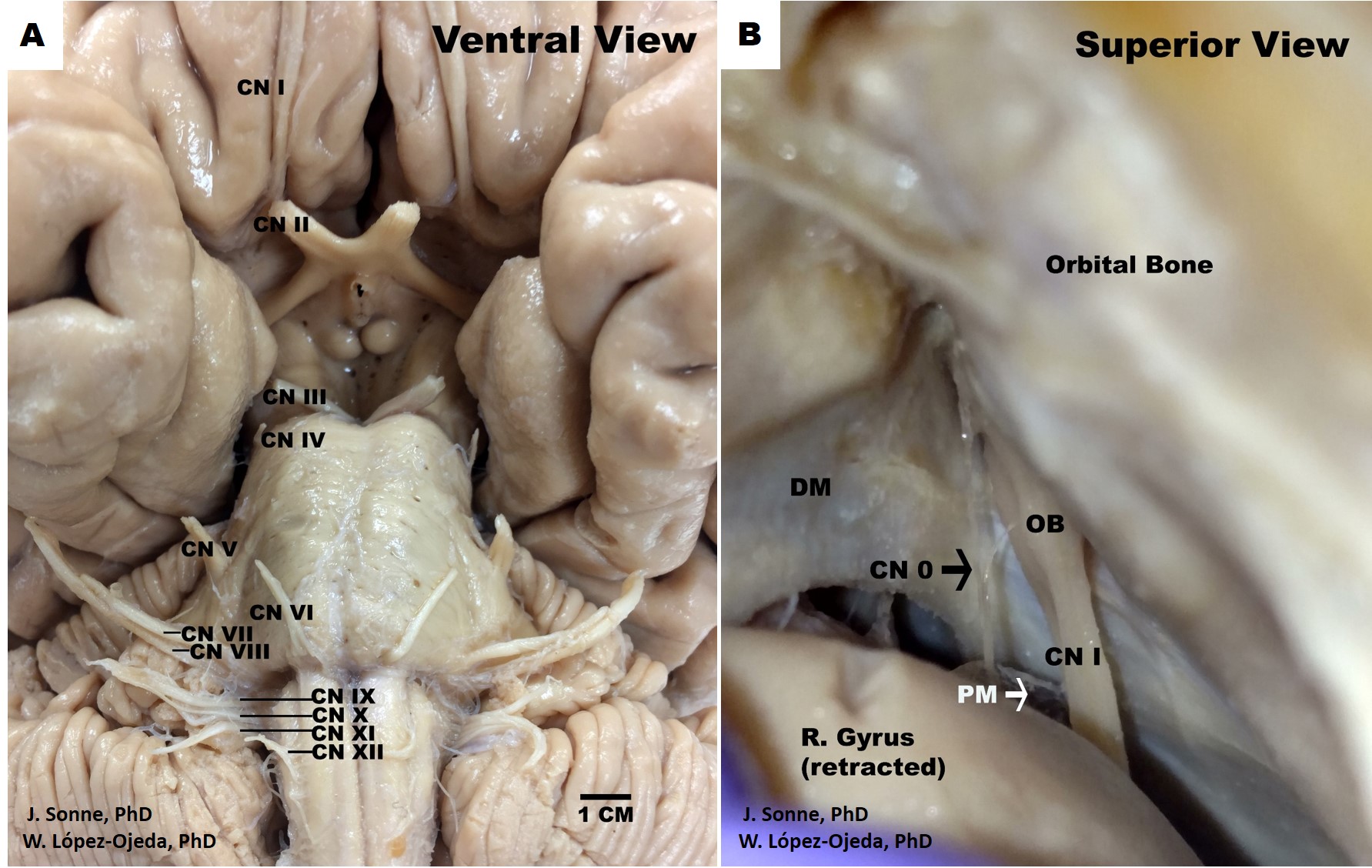
While this is the classical way of organizing and indexing cranial nerves, the scientific reality is more complex and is still debated in the academic realm, including the classification and identification of the routes of distinct cranial nerves fibers and the presence or absence of other less recognized structures, such as the terminal nerve, also called nerve nulla or cranial nerve zero. The latter could be classified as a pure afferent nerve following the traditional organization of cranial nerves—damage to the cranial nerves, their tracts, or nuclei results in stereotyped clinical syndromes.
Structure and Function
The cranial nerves can be considered both in terms of their anatomical numbering from I to XII, which describes their sequential origins from the caudal to the ventral brainstem, or in groups according to their developmental functions (i.e., sensory, motor, mixed). Here we will detail the 12 cranial nerves first broadly in their anatomical ordering before briefly outlining their functional groupings (See Image. Human Brain Cranial Nerves).
Cranial nerve I (olfactory nerve)
Special visceral afferent bipolar sensory neurons reside in the olfactory mucosa, an area of approximately 2 to 4 cm^2 over the superior nasal concha and nasal septum). Here they organize into nerve bundles termed the filia olfactoria, which, together, comprise the olfactory nerve. They enter the cranium through the cribriform plate of the ethmoid bone to synapse in the olfactory bulb, which forms the anterior-most aspect of the olfactory tract at the inferior aspect of the frontal lobe, and which extends posteriorly between the orbital gyrus and the gyrus rectus until bifurcating at the olfactory trigone as the medial and lateral olfactory stria which terminate in higher-order cortex.[2] Some olfactory projections travel medially to the septal area and the contralateral bulb via the anterior commissure, while other fibers travel laterally to the amygdala and piriform cortex, also known as the primary olfactory cortex, where conscious odorant sense is processed.[3]
Cranial nerve II (optic nerve)
Cranial nerve II, the optic nerve, conveys special somatic afferent (SSA) visual sensory information from the rods and cones retinal sensory receptors to the thalamus, especially the lateral geniculate nucleus (LGN) and the superior colliculus (SC). Ganglion cells, whose cell bodies are located deep in the retina, have central projections that form the optic nerve fibers, which traverse the optic canal to enter the cranium. From there, fibers representing the medial visual fields travel posteriorly without crossing at the optic chiasm, while fibers of the lateral visual fields do cross within the chiasm. Therefore, regions of the visual field are retinotopically organized within the optic nerve and at their synapses in the LGN. Collaterals are also given off centrally to innervate the SC, responsible for the pupillary light reflex, and connections within the pulvinar of the thalamus, providing unconscious optic input responsible for the phenomenon of blindsight.[4] In cortically blind individuals, these pulvinar collaterals from the optic nerve allow for unconscious eye movements in response to the detection of light as well as a weak directional sense of the location of the light within the visual field.[5][6][7]
Cranial nerves III, IV, and VI (oculomotor, trochlear, and abducens nerve)
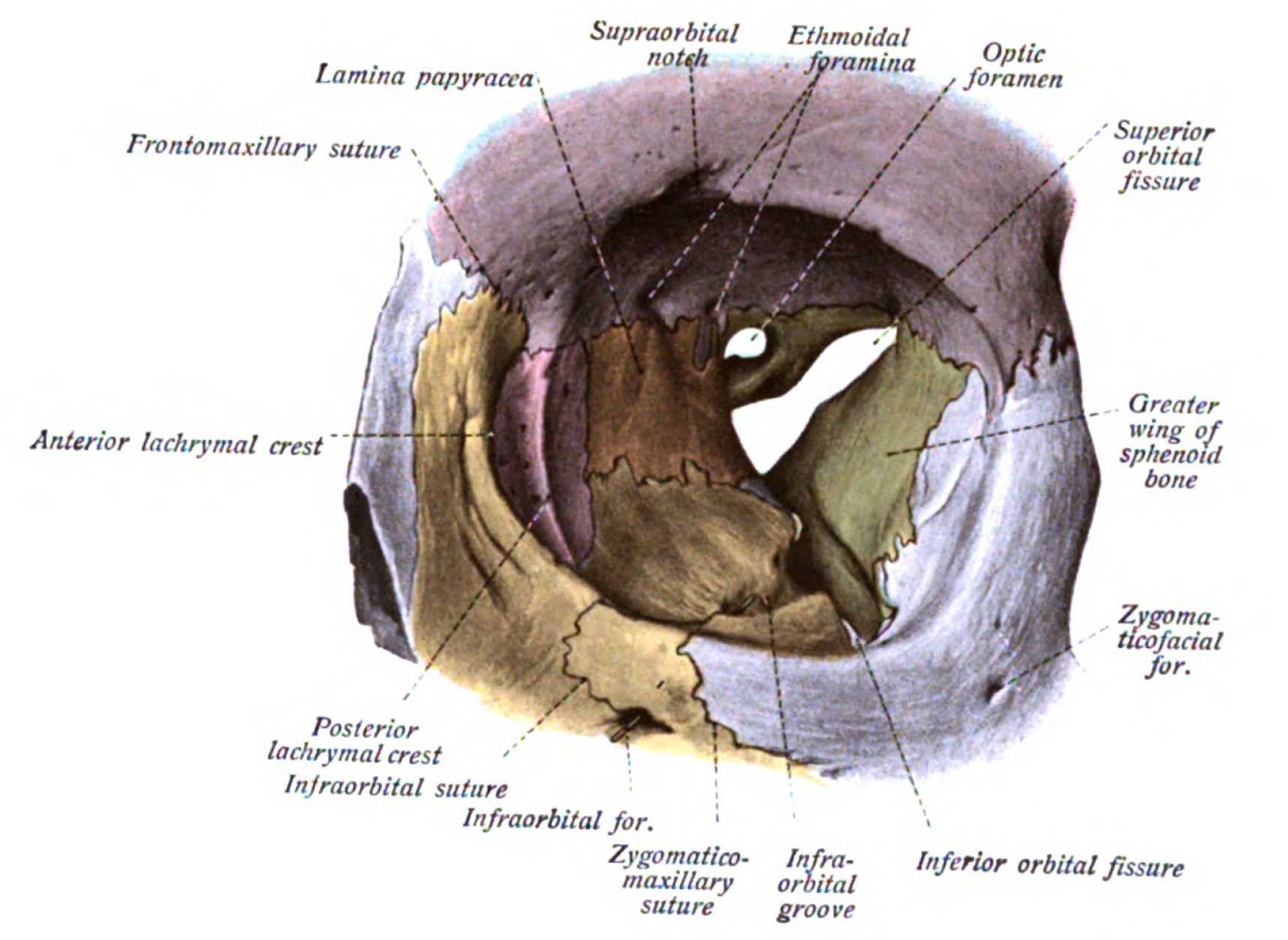
Cranial nerves III, IV, and VI (oculomotor, trochlear, and abducens nerves, respectively) are general somatic efferent (GSE) nerves responsible for innervating the extraocular muscles within the orbit (see Image. Orbit, Anterior View). These nerves travel ipsilaterally from the brainstem to the calvarium through the superior orbital fissure from synonymous brainstem nuclei. The oculomotor nerve (CN III) travels through the common tendinous ring, the common attachment in the posterior orbit for the four extraocular recti muscles, along with the abducens nerve (cranial nerve VI). The trochlear nerve (CN IV) travels into the orbit outside the common tendinous ring to innervate the superior oblique muscle of the eye. The abducens nerve innervates the lateral rectus muscles only; thereby, this nerve can be tested by evaluating the abduction of the eye gaze.
Cranial nerve III innervates most of the eye muscles by splitting into a superior and an inferior branch to innervate the remaining three recti muscles, the inferior oblique, and the skeletal muscle component of the levator palpebrae superioris. However, cranial nerve III also has a general visceral efferent (GVE) component originating in the Edinger-Westphal nucleus (also called accessory oculomotor nucleus or visceral oculomotor nucleus). These fibers travel with cranial nerve III to synapse in the ciliary ganglion within the orbit. The post-ganglionic sympathetic fibers of the ciliary ganglion pierce the sclera of the eye to innervate the pupillary sphincter and ciliary smooth muscles responsible for pupillary constriction and lens accommodation. Pupillary constriction can be tested using the pupillary light reflex through the efferents from the superior colliculus to the accessory oculomotor nucleus.[8]
The eye movements test (abduction, adduction, infraduction, supraduction) effectively assesses the viability of the GSE components of cranial nerves III, IV, and VI.
Cranial nerve V (trigeminal nerve)
Cranial Nerve V is the trigeminal nerve responsible for the general somatic sensory innervation (GSA) of the face through its three main branches, V1, V2, and V3 (ophthalmic, maxillary, and mandibular, respectively). This cranial nerve (via V3) is also responsible for motor innervation (SVE) of the muscles of mastication, the anterior belly of the digastric, mylohyoid, and two tiny tensor muscles: the tensor veli palatini and tensor tympani. While no autonomic fibers travel with the fifth cranial nerve as it exits the pons, parasympathetic fibers from the other mixed cranial nerves will join with peripheral branches of cranial nerve V to innervate their respective target structures, such as the lacrimal, parotid, submandibular and sublingual glands. In this sense, central nuclear or supranuclear lesions may result in ipsilateral sensory or motor deficits, but parasympathetic functions will only be impaired by peripheral nerve damage to the respective branches.[9]
Cranial nerve VII (facial nerve)
Cranial nerve VII (facial nerve) has both motor and autonomic fibers with minor somatosensory components. Special visceral efferent (SVE) motor innervation is to the muscles of facial expression and exits the skull through the stylomastoid foramen deep to the parotid gland. Damage to these fibers results in ipsilateral facial paralysis (facial palsy). General visceral efferents (GVE) and special visceral afferents (SVA) fibers initially exit the brainstem as nervus intermedius, a separate nerve bundle that joins with the other components of the facial nerve within the facial canal. The GVE components from the superior salivary nucleus are responsible for parasympathetic innervation of the glands and mucosae of the face, with the exception of the parotid gland and the smaller buccal and labial glands. Taste fibers from the anterior two-thirds of the tongue travel centrally as the chorda tympani nerve to their cell body of origin in the geniculate ganglion before synapsing centrally in the solitary nucleus. Depending on the location of the lesion, these visceral components may also be impacted in lesions of the facial nerve. Somatic afferents (GSA) provide sensory innervation from the auricle and a small external portion of the auditory canal.[10]
Cranial nerve VIII (vestibulocochlear nerve)
Cranial nerve VIII, the vestibulocochlear nerve, is responsible for the auditory sense and the vestibular sense of orientation of the head. This nerve conveys special sensory afferents (SSA) from the inner ear to the cochlear nuclei and the vestibular nuclei in the caudal medulla oblongata. Hair cells within the cochlear duct, semi-circular canals, utricle, and saccule are polarized sensory receptor cells with apical ciliary extensions that transduce an electrochemical signal upon mechanical deformation. Ganglionic neurons within the cochlea and the vestibular nerve receive this signal peripherally and transmit it centrally through the internal auditory meatus before entering the medulla.[11]
Cranial nerve IX (glossopharyngeal nerve)
Cranial nerve IX (glossopharyngeal nerve) is responsible for motor (SVE) innervation of the stylopharyngeus and the pharyngeal constrictor muscles by the nucleus ambiguus. Inferior salivary nucleus fibers travel with cranial nerve IX to provide general visceral efferent (GVE) innervation to parotid, buccal and labial glands, while visceral afferents (GVA and SVA) receive sensory information from the carotid body and carotid sinus, and taste from the posterior third of the tongue to synapse on the solitary nucleus. The sensory afferents (GSA) receive information from the skin over the tongue, oropharynx, middle ear cavity, and auditory canal.[12]
Cranial nerve X (vagus nerve)
Cranial nerve X is the vagus nerve. The parasympathetic efferents (GVE) fibers from the dorsal vagal nucleus to the thoracic and abdominal viscera to the splenic flexure of the colon represent its major neural component. These fibers form a comprehensive plexus that travels along the esophageal serosa to the viscera. It also has a considerable motor output (SVE) from the nucleus ambiguous to the pharyngeal and soft palate muscles and the intrinsic laryngeal muscles via the superior and recurrent laryngeal nerves. Somatic afferents (GSA) supply the posterior cranial dura and a portion of the ear and external auditory canal epithelium. Visceral afferents (GVA) from the pharynx, larynx, aorta, thoracic and abdominal viscera, and taste buds from the root of the tongue and epiglottis (SVA) synapse on the solitary nucleus as well. Damage to the recurrent laryngeal branch of the vagus nerve can result in vocal hoarseness or acute dyspnea with bilateral avulsion.
Cranial nerve XI (Accessory nerve)
Cranial nerve XI, the spinal accessory nerve, is responsible for the general somatic efferent (GSE) motor innervation of the trapezius and sternocleidomastoid muscles by way of the spinal nucleus of the accessory nerve. The spinal nucleus of the accessory nerve is located within the cervical spinal cord from the levels of C1 through approximately C5/6. The fibers emerge as independent roots, separate from the anterior or dorsal spinal roots of the central spinal grey matter, and ascend through the foramen magnum to enter the cranial cavity. These fibers then exit via the jugular foramen along with cranial nerves IX and X.
Cranial nerve XII (Hypoglossal nerve)
Cranial nerve XII, the hypoglossal nerve, is responsible for the general somatic efferent (GSE) innervation of the intrinsic and extrinsic muscles of the tongue, except the palatoglossus muscle, from the nerve’s synonymous nucleus. This includes the genioglossus, geniohyoid, hyoglossus, and styloglossus muscles. Fibers from the hypoglossal nucleus exit the medulla from the sulcus between the pyramids and the olives as a collection of fibers that coalesce before entering the hypoglossal canal to exit the cranium.[13]
The cranial nerves can also be grouped in terms of their functional roles as efferent nerves (motor output), efferent nerves (sensory input), or mixed nerves. Cranial nerves I, II, and VIII are considered purely afferent nerves since they conduct sensory information from the olfactory region, the retina of the eye, and the inner ear structures, respectively. Cranial nerves III, IV, VI, XI, and XII are considered purely efferent due to their motor output to the orbit, the neck, and the tongue. Cranial nerves V, VII, IX, and X are considered mixed cranial nerves due to the presence of afferent and efferent fibers with both sensory and motor components.
Clinical Significance
CN I
Traumatic injury, especially “whiplash” from automobile collisions, can sever the olfactory projections through the cribriform plate, resulting in anosmia, which has been associated with the development of depression.[14][15][16] The sense of olfaction also appears to have a nonconscious role in activating the limbic system, which may account for such an effect.[17]
CV VIII
Damage to the vestibular component of this nerve causes dizziness, while damage to the cochlear part causes peripheral or sensorineural hearing loss. The internal auditory meatus is a narrow canal of the temporal bone through which these nerves course and schwannoma of the vestibular or cochlear nerves in this meatus easily compresses and impinges these nerves. Early signs and symptoms are progressively worsening hearing loss with tinnitus, and imbalance, leading to a sense of pressure in the ear and facial weakness or paralysis.[18] Vestibular schwannomas have an incidence rate of six to nine new cases per year per million people and are readily treatable with surgery or radiation.[19] However, if the condition is left untreated, it can result in potentially significant and life-threatening tumors.
CN XI
Central root or nuclear damage to the spinal accessory nerve results in ipsilateral flaccid paralysis of the sternocleidomastoid (with difficulty turning the head against force) and partial ipsilateral trapezius paralysis leading to shoulder drop. The trapezius is innervated by anterior horn grey matter from cervical spinal regions C3 through C4/5 in addition to the spinal accessory nerve. Thus complete paralysis of the trapezius muscle will not occur following a simple focal lesion.[20]
CN XII
Damage to the nucleus or nerve fibers results in tongue deviation toward the side of the lesion, as the ipsilateral genioglossus muscle becomes weak or flaccid, reducing its ability to protrude the tongue.
Other Issues
In addition, the terminal nerve or cranial nerve zero has been identified as a separate cranial nerve in the human brain dating back to 1914 but is overlooked by most current anatomical textbooks.[21] Also called nervus terminalis (for its proximity to the lamina terminalis) or the nerve nulla (i.e., nothing, zero), cranial nerve zero is comprised of an independent central plexus of small unmyelinated (possibly special visceral afferent [SVA]) fibers located medially and in very close proximity to the olfactory tract by the olfactory trigone. Its discrete positioning may explain its poor identification during standard dissection techniques. The cranial nerve zero fibers travel centrally to subcortical structures; it sends projections to the medial pre-commissural septum and the medial septal nucleus, among others. It appears to have a rich bundle of well-vascularized fibers ascending from the nasal submucosa and projecting to important limbic structures (e.g., amygdala, hypothalamic nuclei).
Functionally it has been regarded as pheromones processing unconsciously by regulating autonomic responses through hypothalamic gonadotropin-releasing hormone (GnRH), possibly via the kisspeptin neuronal network. Clinically, a disruption of the normal embryological migratory pathway of GnRH neural crest cells of the olfactory placode and basal forebrain can result in Kallman syndrome, a genetic condition characterized by hypogonadotropic hypogonadism with partial or total anosmia, also resulting in abnormal sexual development in both sexes.[22]
The canonical descriptions of the cranial nerves serve as an overview of the current medical literature, but research advances continuously reveal new information leading to riveting debates regarding the original classification of some cranial nerve fibers. For instance, it has been found that the nervus intermedius, known for carrying taste fibers from the tongue and associated with cranial nerve VII, has visceral efferent connections with the vestibulocochlear nerve in the internal auditory meatus.[23] Indeed, varied populations of efferent fibers to the organ of Corti hair cells modulate afferent transduction sensitivity and are present in mammals and vertebrates.[24][25][26][27][25]
Also, the classification of the cranial accessory nerve as a portion of either vagus or accessory nerves is continually disputed. In the majority of cases cranial accessory nerve, with its fibers originating in the nucleus ambiguus of the medulla, has no connections with the spinal accessory nerve and its fibers originating in the spinal accessory nucleus of the spinal cord.[28] Also, cranial nerve zero has been recognized in human embryos since 1905 and in adult humans since 1914. While historical textbooks included this nerve, modern anatomical textbooks have omitted it from the official list of cranial nerves.[29][30]
However, the sensory function and modulation of the hypothalamic endocrine and autonomic systems make it physiologically and clinically relevant.[21] Similarly, it has been argued that the optic nerve (cranial nerve II) is not a peripheral nerve but a central cranial tract.[29] This is evidence of the continuous changes brought to accepted anatomical and medical terminology and the structures reflected by those terms.[31][32] As such, a holistic view of cranial nerves may be more valuable than the categorical organization currently present in most textbook narratives. These issues warrant attention since all cranial nerve nuances are particularly relevant for medical and health professions education.