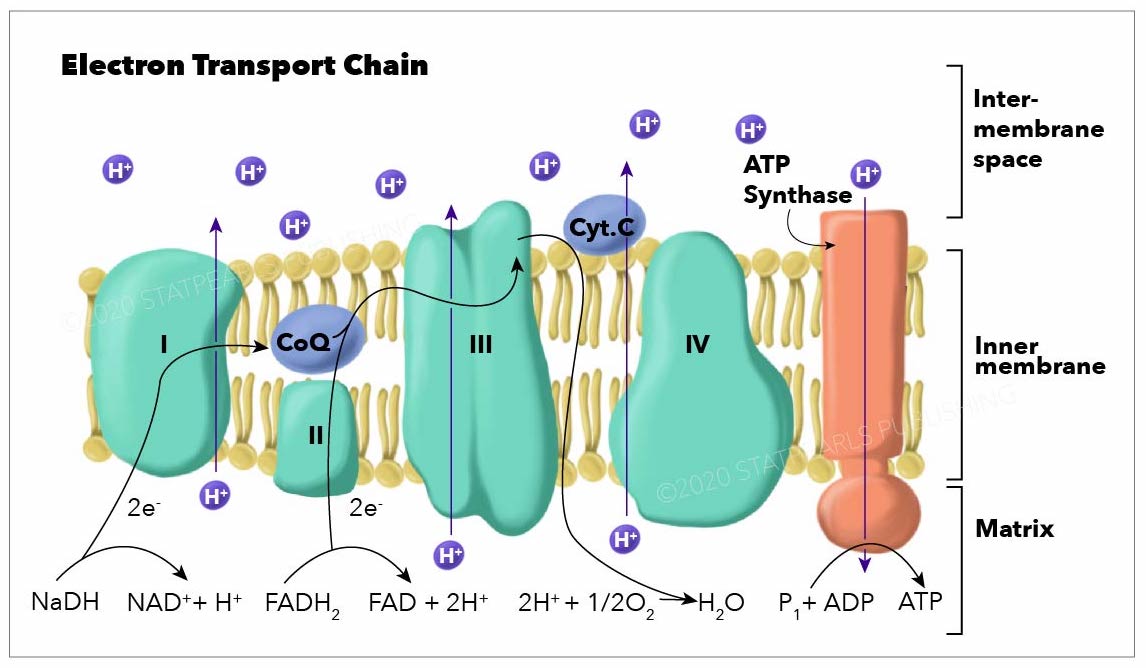
[1]
Lencina AM, Franza T, Sullivan MJ, Ulett GC, Ipe DS, Gaudu P, Gennis RB, Schurig-Briccio LA. Type 2 NADH Dehydrogenase Is the Only Point of Entry for Electrons into the Streptococcus agalactiae Respiratory Chain and Is a Potential Drug Target. mBio. 2018 Jul 3:9(4):. doi: 10.1128/mBio.01034-18. Epub 2018 Jul 3
[PubMed PMID: 29970468]
[2]
Hirst J. Towards the molecular mechanism of respiratory complex I. The Biochemical journal. 2009 Dec 23:425(2):327-39. doi: 10.1042/BJ20091382. Epub 2009 Dec 23
[PubMed PMID: 20025615]
[3]
Sazanov LA, Hinchliffe P. Structure of the hydrophilic domain of respiratory complex I from Thermus thermophilus. Science (New York, N.Y.). 2006 Mar 10:311(5766):1430-6
[PubMed PMID: 16469879]
[4]
Hirst J. Energy transduction by respiratory complex I--an evaluation of current knowledge. Biochemical Society transactions. 2005 Jun:33(Pt 3):525-9
[PubMed PMID: 15916556]
[5]
Yankovskaya V, Horsefield R, Törnroth S, Luna-Chavez C, Miyoshi H, Léger C, Byrne B, Cecchini G, Iwata S. Architecture of succinate dehydrogenase and reactive oxygen species generation. Science (New York, N.Y.). 2003 Jan 31:299(5607):700-4
[PubMed PMID: 12560550]
[6]
Horsefield R, Iwata S, Byrne B. Complex II from a structural perspective. Current protein & peptide science. 2004 Apr:5(2):107-18
[PubMed PMID: 15078221]
Level 3 (low-level) evidence
[7]
Geertman JM, van Maris AJ, van Dijken JP, Pronk JT. Physiological and genetic engineering of cytosolic redox metabolism in Saccharomyces cerevisiae for improved glycerol production. Metabolic engineering. 2006 Nov:8(6):532-42
[PubMed PMID: 16891140]
[8]
Thorpe C, Kim JJ. Structure and mechanism of action of the acyl-CoA dehydrogenases. FASEB journal : official publication of the Federation of American Societies for Experimental Biology. 1995 Jun:9(9):718-25
[PubMed PMID: 7601336]
[9]
Sun C, Benlekbir S, Venkatakrishnan P, Wang Y, Hong S, Hosler J, Tajkhorshid E, Rubinstein JL, Gennis RB. Structure of the alternative complex III in a supercomplex with cytochrome oxidase. Nature. 2018 May:557(7703):123-126. doi: 10.1038/s41586-018-0061-y. Epub 2018 Apr 25
[PubMed PMID: 29695868]
[10]
Iwata S, Lee JW, Okada K, Lee JK, Iwata M, Rasmussen B, Link TA, Ramaswamy S, Jap BK. Complete structure of the 11-subunit bovine mitochondrial cytochrome bc1 complex. Science (New York, N.Y.). 1998 Jul 3:281(5373):64-71
[PubMed PMID: 9651245]
[11]
Trumpower BL. The protonmotive Q cycle. Energy transduction by coupling of proton translocation to electron transfer by the cytochrome bc1 complex. The Journal of biological chemistry. 1990 Jul 15:265(20):11409-12
[PubMed PMID: 2164001]
[12]
Hunte C, Palsdottir H, Trumpower BL. Protonmotive pathways and mechanisms in the cytochrome bc1 complex. FEBS letters. 2003 Jun 12:545(1):39-46
[PubMed PMID: 12788490]
Level 3 (low-level) evidence
[13]
Calhoun MW, Thomas JW, Gennis RB. The cytochrome oxidase superfamily of redox-driven proton pumps. Trends in biochemical sciences. 1994 Aug:19(8):325-30
[PubMed PMID: 7940677]
[14]
Schmidt-Rohr K. Oxygen Is the High-Energy Molecule Powering Complex Multicellular Life: Fundamental Corrections to Traditional Bioenergetics. ACS omega. 2020 Feb 11:5(5):2221-2233. doi: 10.1021/acsomega.9b03352. Epub 2020 Jan 28
[PubMed PMID: 32064383]
[15]
Lovero D, Giordano L, Marsano RM, Sanchez-Martinez A, Boukhatmi H, Drechsler M, Oliva M, Whitworth AJ, Porcelli D, Caggese C. Characterization of Drosophila ATPsynC mutants as a new model of mitochondrial ATP synthase disorders. PloS one. 2018:13(8):e0201811. doi: 10.1371/journal.pone.0201811. Epub 2018 Aug 10
[PubMed PMID: 30096161]
[16]
Okuno D, Iino R, Noji H. Rotation and structure of FoF1-ATP synthase. Journal of biochemistry. 2011 Jun:149(6):655-64. doi: 10.1093/jb/mvr049. Epub 2011 Apr 26
[PubMed PMID: 21524994]
[17]
Junge W, Nelson N. ATP synthase. Annual review of biochemistry. 2015:84():631-57. doi: 10.1146/annurev-biochem-060614-034124. Epub 2015 Mar 23
[PubMed PMID: 25839341]
[18]
Hinkle PC. P/O ratios of mitochondrial oxidative phosphorylation. Biochimica et biophysica acta. 2005 Jan 7:1706(1-2):1-11
[PubMed PMID: 15620362]
[19]
Barrett MA, Zheng S, Roshankar G, Alsop RJ, Belanger RK, Huynh C, Kučerka N, Rheinstädter MC. Interaction of aspirin (acetylsalicylic acid) with lipid membranes. PloS one. 2012:7(4):e34357. doi: 10.1371/journal.pone.0034357. Epub 2012 Apr 17
[PubMed PMID: 22529913]
[20]
Warrick BJ, King A, Smolinske S, Thomas R, Aaron C. A 29-year analysis of acute peak salicylate concentrations in fatalities reported to United States poison centers. Clinical toxicology (Philadelphia, Pa.). 2018 Sep:56(9):846-851. doi: 10.1080/15563650.2018.1435887. Epub 2018 Feb 12
[PubMed PMID: 29431532]
[22]
Enerbäck S. The origins of brown adipose tissue. The New England journal of medicine. 2009 May 7:360(19):2021-3. doi: 10.1056/NEJMcibr0809610. Epub
[PubMed PMID: 19420373]
[23]
Zhou W, Faraldo-Gómez JD. Membrane plasticity facilitates recognition of the inhibitor oligomycin by the mitochondrial ATP synthase rotor. Biochimica et biophysica acta. Bioenergetics. 2018 Sep:1859(9):789-796. doi: 10.1016/j.bbabio.2018.03.019. Epub 2018 Apr 7
[PubMed PMID: 29630891]
[24]
Kamalian L, Douglas O, Jolly CE, Snoeys J, Simic D, Monshouwer M, Williams DP, Kevin Park B, Chadwick AE. The utility of HepaRG cells for bioenergetic investigation and detection of drug-induced mitochondrial toxicity. Toxicology in vitro : an international journal published in association with BIBRA. 2018 Dec:53():136-147. doi: 10.1016/j.tiv.2018.08.001. Epub 2018 Aug 7
[PubMed PMID: 30096366]
[25]
Wood DM, Alsahaf H, Streete P, Dargan PI, Jones AL. Fatality after deliberate ingestion of the pesticide rotenone: a case report. Critical care (London, England). 2005 Jun:9(3):R280-4
[PubMed PMID: 15987402]
Level 3 (low-level) evidence
[26]
Lupescu A, Jilani K, Zbidah M, Lang F. Induction of apoptotic erythrocyte death by rotenone. Toxicology. 2012 Oct 28:300(3):132-7. doi: 10.1016/j.tox.2012.06.007. Epub 2012 Jun 19
[PubMed PMID: 22727881]
[27]
Wallace KB. Doxorubicin-induced cardiac mitochondrionopathy. Pharmacology & toxicology. 2003 Sep:93(3):105-15
[PubMed PMID: 12969434]
[28]
Weaver LK. Clinical practice. Carbon monoxide poisoning. The New England journal of medicine. 2009 Mar 19:360(12):1217-25. doi: 10.1056/NEJMcp0808891. Epub
[PubMed PMID: 19297574]
[29]
Sato K, Tamaki K, Hattori H, Moore CM, Tsutsumi H, Okajima H, Katsumata Y. Determination of total hemoglobin in forensic blood samples with special reference to carboxyhemoglobin analysis. Forensic science international. 1990 Nov:48(1):89-96
[PubMed PMID: 2279722]
[30]
Barker SJ, Tremper KK. The effect of carbon monoxide inhalation on pulse oximetry and transcutaneous PO2. Anesthesiology. 1987 May:66(5):677-9
[PubMed PMID: 3578881]
[31]
Raub JA, Mathieu-Nolf M, Hampson NB, Thom SR. Carbon monoxide poisoning--a public health perspective. Toxicology. 2000 Apr 7:145(1):1-14
[PubMed PMID: 10771127]
Level 3 (low-level) evidence
[32]
Jensen P, Wilson MT, Aasa R, Malmström BG. Cyanide inhibition of cytochrome c oxidase. A rapid-freeze e.p.r. investigation. The Biochemical journal. 1984 Dec 15:224(3):829-37
[PubMed PMID: 6098268]
[33]
Shchepina LA, Pletjushkina OY, Avetisyan AV, Bakeeva LE, Fetisova EK, Izyumov DS, Saprunova VB, Vyssokikh MY, Chernyak BV, Skulachev VP. Oligomycin, inhibitor of the F0 part of H+-ATP-synthase, suppresses the TNF-induced apoptosis. Oncogene. 2002 Nov 21:21(53):8149-57
[PubMed PMID: 12444550]