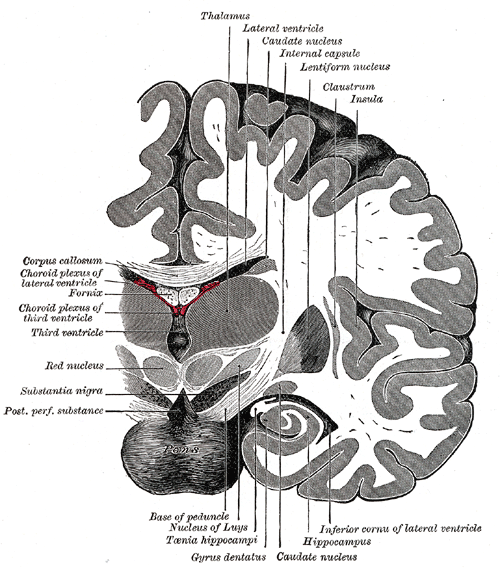
[1]
Grahn JA, Parkinson JA, Owen AM. The cognitive functions of the caudate nucleus. Progress in neurobiology. 2008 Nov:86(3):141-55. doi: 10.1016/j.pneurobio.2008.09.004. Epub 2008 Sep 7
[PubMed PMID: 18824075]
[2]
Fisher HE, Aron A, Brown LL. Romantic love: a mammalian brain system for mate choice. Philosophical transactions of the Royal Society of London. Series B, Biological sciences. 2006 Dec 29:361(1476):2173-86
[PubMed PMID: 17118931]
[3]
Kotz SA, Anwander A, Axer H, Knösche TR. Beyond cytoarchitectonics: the internal and external connectivity structure of the caudate nucleus. PloS one. 2013:8(7):e70141. doi: 10.1371/journal.pone.0070141. Epub 2013 Jul 26
[PubMed PMID: 23922939]
[4]
Pellizzaro Venti M, Paciaroni M, Caso V. Caudate infarcts and hemorrhages. Frontiers of neurology and neuroscience. 2012:30():137-40. doi: 10.1159/000333616. Epub 2012 Feb 14
[PubMed PMID: 22377881]
[5]
Hikosaka O, Takikawa Y, Kawagoe R. Role of the basal ganglia in the control of purposive saccadic eye movements. Physiological reviews. 2000 Jul:80(3):953-78
[PubMed PMID: 10893428]
[6]
Graff-Radford J, Williams L, Jones DT, Benarroch EE. Caudate nucleus as a component of networks controlling behavior. Neurology. 2017 Nov 21:89(21):2192-2197. doi: 10.1212/WNL.0000000000004680. Epub 2017 Oct 25
[PubMed PMID: 29070661]
[7]
Seger CA, Cincotta CM. The roles of the caudate nucleus in human classification learning. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2005 Mar 16:25(11):2941-51
[PubMed PMID: 15772354]
[8]
Barkóczi B, Nagypál T, Nyujtó D, Katona X, Eördegh G, Bodosi B, Benedek G, Braunitzer G, Nagy A. Background activity and visual responsiveness of caudate nucleus neurons in halothane anesthetized and in awake, behaving cats. Neuroscience. 2017 Jul 25:356():182-192. doi: 10.1016/j.neuroscience.2017.05.028. Epub 2017 May 22
[PubMed PMID: 28546109]
[9]
White NM. Some highlights of research on the effects of caudate nucleus lesions over the past 200 years. Behavioural brain research. 2009 Apr 12:199(1):3-23. doi: 10.1016/j.bbr.2008.12.003. Epub 2008 Dec 7
[PubMed PMID: 19111791]
[10]
Yanike M, Ferrera VP. Representation of outcome risk and action in the anterior caudate nucleus. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2014 Feb 26:34(9):3279-90. doi: 10.1523/JNEUROSCI.3818-13.2014. Epub
[PubMed PMID: 24573287]
[11]
Müller NCJ, Konrad BN, Kohn N, Muñoz-López M, Czisch M, Fernández G, Dresler M. Hippocampal-caudate nucleus interactions support exceptional memory performance. Brain structure & function. 2018 Apr:223(3):1379-1389. doi: 10.1007/s00429-017-1556-2. Epub 2017 Nov 14
[PubMed PMID: 29138923]
[12]
Hasegawa H, Matsuura M, Sato H, Yamamoto T, Kanai H. Imaging of gaps in digital joints by measurement of ultrasound transmission using a linear array. Ultrasound in medicine & biology. 2009 Mar:35(3):382-94. doi: 10.1016/j.ultrasmedbio.2008.09.022. Epub 2008 Dec 27
[PubMed PMID: 19111971]
[14]
Onorati M, Castiglioni V, Biasci D, Cesana E, Menon R, Vuono R, Talpo F, Laguna Goya R, Lyons PA, Bulfamante GP, Muzio L, Martino G, Toselli M, Farina C, Barker RA, Biella G, Cattaneo E. Molecular and functional definition of the developing human striatum. Nature neuroscience. 2014 Dec:17(12):1804-15. doi: 10.1038/nn.3860. Epub 2014 Nov 10
[PubMed PMID: 25383901]
[15]
Dayer AG, Cleaver KM, Abouantoun T, Cameron HA. New GABAergic interneurons in the adult neocortex and striatum are generated from different precursors. The Journal of cell biology. 2005 Jan 31:168(3):415-27
[PubMed PMID: 15684031]
[16]
Luzzati F, De Marchis S, Fasolo A, Peretto P. Neurogenesis in the caudate nucleus of the adult rabbit. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2006 Jan 11:26(2):609-21
[PubMed PMID: 16407559]
[17]
Ernst A, Alkass K, Bernard S, Salehpour M, Perl S, Tisdale J, Possnert G, Druid H, Frisén J. Neurogenesis in the striatum of the adult human brain. Cell. 2014 Feb 27:156(5):1072-83. doi: 10.1016/j.cell.2014.01.044. Epub 2014 Feb 20
[PubMed PMID: 24561062]
[18]
Tatu L, Moulin T, Bogousslavsky J, Duvernoy H. Arterial territories of the human brain: cerebral hemispheres. Neurology. 1998 Jun:50(6):1699-708
[PubMed PMID: 9633714]
[19]
Feekes JA, Cassell MD. The vascular supply of the functional compartments of the human striatum. Brain : a journal of neurology. 2006 Aug:129(Pt 8):2189-201
[PubMed PMID: 16815876]
[20]
Plog BA, Nedergaard M. The Glymphatic System in Central Nervous System Health and Disease: Past, Present, and Future. Annual review of pathology. 2018 Jan 24:13():379-394. doi: 10.1146/annurev-pathol-051217-111018. Epub
[PubMed PMID: 29195051]
[21]
Rijpkema M, Everaerd D, van der Pol C, Franke B, Tendolkar I, Fernández G. Normal sexual dimorphism in the human basal ganglia. Human brain mapping. 2012 May:33(5):1246-52. doi: 10.1002/hbm.21283. Epub 2011 Apr 26
[PubMed PMID: 21523857]
[22]
Lenroot RK, Gogtay N, Greenstein DK, Wells EM, Wallace GL, Clasen LS, Blumenthal JD, Lerch J, Zijdenbos AP, Evans AC, Thompson PM, Giedd JN. Sexual dimorphism of brain developmental trajectories during childhood and adolescence. NeuroImage. 2007 Jul 15:36(4):1065-73
[PubMed PMID: 17513132]
[23]
Hervais-Adelman A, Egorova N, Golestani N. Beyond bilingualism: multilingual experience correlates with caudate volume. Brain structure & function. 2018 Sep:223(7):3495-3502. doi: 10.1007/s00429-018-1695-0. Epub 2018 Jun 14
[PubMed PMID: 29948191]
[24]
Isaacs EB, Gadian DG, Sabatini S, Chong WK, Quinn BT, Fischl BR, Lucas A. The effect of early human diet on caudate volumes and IQ. Pediatric research. 2008 Mar:63(3):308-14. doi: 10.1203/PDR.0b013e318163a271. Epub
[PubMed PMID: 18287970]
[25]
Willford J,Day R,Aizenstein H,Day N, Caudate asymmetry: a neurobiological marker of moderate prenatal alcohol exposure in young adults. Neurotoxicology and teratology. 2010 Nov-Dec;
[PubMed PMID: 20609385]
[26]
Warton FL, Meintjes EM, Warton CMR, Molteno CD, Lindinger NM, Carter RC, Zöllei L, Wintermark P, Jacobson JL, van der Kouwe A, Jacobson SW. Prenatal methamphetamine exposure is associated with reduced subcortical volumes in neonates. Neurotoxicology and teratology. 2018 Jan-Feb:65():51-59. doi: 10.1016/j.ntt.2017.10.005. Epub 2017 Oct 22
[PubMed PMID: 29069607]
[27]
Aouizerate B,Cuny E,Martin-Guehl C,Guehl D,Amieva H,Benazzouz A,Fabrigoule C,Allard M,Rougier A,Bioulac B,Tignol J,Burbaud P, Deep brain stimulation of the ventral caudate nucleus in the treatment of obsessive-compulsive disorder and major depression. Case report. Journal of neurosurgery. 2004 Oct;
[PubMed PMID: 15481726]
Level 3 (low-level) evidence
[29]
Im I, Jun JP, Hwang S, Ko MH. Swallowing outcomes in patients with subcortical stroke associated with lesions of the caudate nucleus and insula. The Journal of international medical research. 2018 Sep:46(9):3552-3562. doi: 10.1177/0300060518775290. Epub 2018 Jun 4
[PubMed PMID: 29865925]
[30]
Ances BM,Roc AC,Wang J,Korczykowski M,Okawa J,Stern J,Kim J,Wolf R,Lawler K,Kolson DL,Detre JA, Caudate blood flow and volume are reduced in HIV neurocognitively impaired patients. Neurology. 2006 Mar 28;
[PubMed PMID: 16567703]
[31]
Looi JC, Lindberg O, Zandbelt BB, Ostberg P, Andersen C, Botes L, Svensson L, Wahlund LO. Caudate nucleus volumes in frontotemporal lobar degeneration: differential atrophy in subtypes. AJNR. American journal of neuroradiology. 2008 Sep:29(8):1537-43. doi: 10.3174/ajnr.A1168. Epub
[PubMed PMID: 18782907]
[32]
Blumberg HP, Stern E, Martinez D, Ricketts S, de Asis J, White T, Epstein J, McBride PA, Eidelberg D, Kocsis JH, Silbersweig DA. Increased anterior cingulate and caudate activity in bipolar mania. Biological psychiatry. 2000 Dec 1:48(11):1045-52
[PubMed PMID: 11094137]
[33]
Narayanaswamy JC,Jose DA,Kalmady SV,Venkatasubramanian G,Janardhana Reddy YC, Clinical correlates of caudate volume in drug-naïve adult patients with obsessive-compulsive disorder. Psychiatry research. 2013 Apr 30;
[PubMed PMID: 23489673]
[34]
Waltzman D, Soman S, Hantke NC, Fairchild JK, Kinoshita LM, Wintermark M, Ashford JW, Yesavage J, Williams L, Adamson MM, Furst AJ. Altered Microstructural Caudate Integrity in Posttraumatic Stress Disorder but Not Traumatic Brain Injury. PloS one. 2017:12(1):e0170564. doi: 10.1371/journal.pone.0170564. Epub 2017 Jan 23
[PubMed PMID: 28114393]
[35]
Tremols V, Bielsa A, Soliva JC, Raheb C, Carmona S, Tomas J, Gispert JD, Rovira M, Fauquet J, Tobeña A, Bulbena A, Vilarroya O. Differential abnormalities of the head and body of the caudate nucleus in attention deficit-hyperactivity disorder. Psychiatry research. 2008 Aug 30:163(3):270-8. doi: 10.1016/j.pscychresns.2007.04.017. Epub 2008 Jul 24
[PubMed PMID: 18656331]
[36]
Okugawa G, Nobuhara K, Takase K, Saito Y, Yoshimura M, Kinoshita T. Olanzapine increases grey and white matter volumes in the caudate nucleus of patients with schizophrenia. Neuropsychobiology. 2007:55(1):43-6
[PubMed PMID: 17556852]
[37]
Langen M, Durston S, Staal WG, Palmen SJ, van Engeland H. Caudate nucleus is enlarged in high-functioning medication-naive subjects with autism. Biological psychiatry. 2007 Aug 1:62(3):262-6
[PubMed PMID: 17224135]
[38]
Naismith S, Hickie I, Ward PB, Turner K, Scott E, Little C, Mitchell P, Wilhelm K, Parker G. Caudate nucleus volumes and genetic determinants of homocysteine metabolism in the prediction of psychomotor speed in older persons with depression. The American journal of psychiatry. 2002 Dec:159(12):2096-8
[PubMed PMID: 12450963]
[39]
Foundas AL, Mock JR, Cindass R Jr, Corey DM. Atypical caudate anatomy in children who stutter. Perceptual and motor skills. 2013 Apr:116(2):528-43
[PubMed PMID: 24032328]
[40]
Henkel K, Danek A, Grafman J, Butman J, Kassubek J. Head of the caudate nucleus is most vulnerable in chorea-acanthocytosis: a voxel-based morphometry study. Movement disorders : official journal of the Movement Disorder Society. 2006 Oct:21(10):1728-31
[PubMed PMID: 16874760]
[41]
Seghatoleslam M, Ghadiri MK, Ghaffarian N, Speckmann EJ, Gorji A. Cortical spreading depression modulates the caudate nucleus activity. Neuroscience. 2014 May 16:267():83-90. doi: 10.1016/j.neuroscience.2014.02.029. Epub 2014 Mar 6
[PubMed PMID: 24613721]