Introduction
Astrocytes are a subtype of glial cells that make up the majority of cells in the human central nervous system (CNS). They perform metabolic, structural, homeostatic, and neuroprotective tasks such as clearing excess neurotransmitters, stabilizing and regulating the blood-brain barrier, and promoting synapse formation.[1][2][3][4] Unlike neurons and other cells in the nervous system, they do not conduct electrical signals. Instead, they ensure the continued function of their signal-relaying counterparts.[1] Due to their diverse array of responsibilities, science has viewed as both targets for treatment and causes of disease. Thus, it is crucial for clinicians to understand the histology and purpose of astrocytes, and to identify their roles in both healthy and pathological states.
Issues of Concern
The broad and extensive involvement of astrocytes in many aspects of CNS development and function suggests that they carry implications in an array of different nervous system disorders. Specifically, any condition or stimulus that would impair the ability of astrocytes to function would effectively disturb the blood-brain barrier, synapse formation and stabilization, signaling efficiency, energy metabolism, and fluid-ion homeostasis in the CNS.[5] Deficiencies in these functions could potentially cause serious damage to the CNS.
Structure
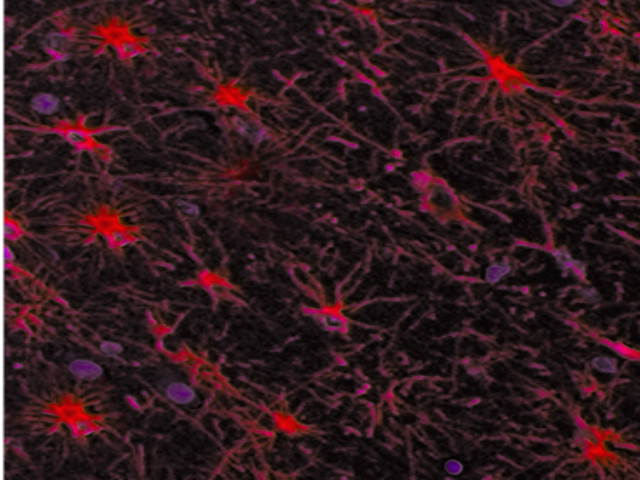
Astrocytes derive their name from their radially-arranged foot processes that give them a star-like appearance. These foot processes, which are marked by a protein known as glial fibrillary acidic protein (GFAP), allow astrocytes to communicate with and affect surrounding neural vasculature. Astrocytes also couple with each other through gap junctions, forming a network that coordinates their responses to stress and other stimuli.[1][2] In the human nervous system, there are four main types of astrocytes: interlaminar, protoplasmic, varicose projection, and fibrous.[1]
Interlaminar astrocytes, which are unique to humans and other primates, possess radial tangential fibers and vertical “cable-like” processes.[1] These astrocytes are so named because, although their cell bodies originate in layer I, they project their long vertical processes, which measure up to one millimeter, to layers III and IV of the cerebral cortex.[1][2] They have small cell bodies, exhibit a high density of GFAP fibers, and exist in abundance in the human CNS.[1]
Protoplasmic astrocytes make up the majority of astrocytes. They are large astrocytes with a high density of tortuous, bulbous GFAP-positive processes that branch into smaller, finer processes.[2] Distributed uniformly throughout the grey matter of cortical layers II through VI, protoplasmic astrocytes organize themselves into separate domains in which each astrocyte projects to the surrounding vasculature and synapses with little overlap.[2] Current conjecture regarding this domain organization suggests that, because of their pervasive projections, protoplasmic astrocytes may modulate and synchronize blood flow with synaptic activity to improve neural efficiency.[2]
In contrast to the protoplasmic astroglia, fibrous astroglia reside around white matter tracts.[1][2]These cells are larger than protoplasmic astrocytes, but they express fewer foot processes than other astrocyte types.[1][2] Since white matter tracts have few synapses within them, suspicions are that fibrous astrocytes do not regulate synaptic activity and instead communicate almost exclusively with the neurovasculature to regulate metabolic and ionic homeostasis in the CNS.[1]
The last major astrocyte phenotype in the human CNS is the varicose projection astroglia. These astrocytes occupy cortical layers V and VI and, like interlaminar astrocytes, have long foot processes that may connect with cells in other cortical layers.[1] These processes, which range in number from one to five, measure up to one millimeter in length, with spiny branches and varicosities every 10 micrometers.[1][2] Although their specific function is unknown, the thought is that interlaminar and varicose projection astrocytes allow for higher-order cognitive processing because only primates' nervous systems contain these specific astroglial types.[2]
Function
Although research has yet to determine specific tasks unique to each astrocyte type, scientists widely accept that astrocytes play a pivotal role in structural support, metabolic regulation, neuroprotection, growth, and homeostatic maintenance of the central nervous system.[3][4][6][7] To fulfill the responsibilities demanded of them, astrocytes are equipped with gap junctions, ion channels, and foot processes that connect them with neurons and their vascular supply within the CNS, allowing astrocytes to monitor and interact with the extracellular environment as well as the neural network. The gap junctions formed between astrocytes, which contain connexins 30 and 43, enable inter-astrocyte communication via glial transmission using calcium signals.[1][4] In this way, the foot processes and ion channels enable astrocytes to regulate the CNS environment.[1][4]
Using these tools, astrocytes perform a number of specific functions: neurotransmitter uptake and recycling, promotion of neuronal survival and synapse formation, regeneration of CNS damage, propagation of signal transmission between neurons via calcium signaling, potassium ion buffering, maintenance of pH and fluid levels in the brain, removal of free radicals in the CNS, regulation of the blood-brain barrier, coordination of cerebral blood flow based on regional neuronal needs, and secretion of trophic factors to guide axons and to promote development of neurons.[1][3][4][6] Future research may show that astrocytes are capable of even more functions than these.[3][4]
Metabolically, astrocytes coordinate cerebral blood flow based on regional neuronal needs. They synthesize glycogen, provide glycolytic components for neurons during periods of high demand, and remove free radicals from the CNS.[3][4] Since astrocytes directly connect to the neurovasculature through their foot processes and can transmit as well as receive complex signals via calcium signaling, astrocytes are in a prime position to elicit vasodilation or vasoconstriction to adjust nutrient-waste exchange for surrounding neurons based on their activity.[4] Astrocytes can then provide further metabolic support to the CNS by taking in glucose via GLUT-1 and synthesizing glycogen when stimulated by insulin or L-glutamate, which allows them to store energy for situations when glucose is low.[4] When neurons need additional energy, astrocytes can then tap into these glycogen stores by synthesizing lactic acid and sending it to neurons via monocarboxylate transporters (MCTs) 1 and 4, which in turn take in the lactic acid for energy via MCT2.[4] When metabolic reactions fueling the CNS result in excessive free radicals, astrocytes use a cystine-glutamate exchanger to store cystine and protect neurons from oxidative damage. Astrocytes can then convert cystine into cysteine which then converts into the antioxidant glutathione, a substance released into the extracellular space in precursor form for neurons to use.[4]
Astrocytes maintain CNS homeostasis by regulating pH and fluid levels in the brain, buffering potassium ion concentration in the extracellular fluid, taking up and recycling excess neurotransmitters, and detoxifying ammonia.[3][4] Astrocytes possess a supply of carbonic anhydrase that enables them to convert waste CO2 from neurons into bicarbonate. Astrocytes use their MCTs to co-transport H+ and lactate for metabolism.[4] These two mechanisms are thought to allow astrocytes to modulate the acid-base balance of the extracellular matrix. Astrocytes also possess a Kir4.1 potassium channel transporter as well as aquaporin 4 (AQP4).[4][5] These transporters are expressed together on astrocyte foot processes to regulate water volume in the extracellular matrix as well as cellular potential. Astrocytes can thus control action potential firing.[4][5] Perhaps the best-known homeostatic function of astrocytes is glutamate and GABA uptake and recycling into glutamine. Astrocytes use ATP and the Na+ gradient created by surrounding neurons to take in glutamate, along with 3 Na+, 1 Cl-, and 1 H+, in exchange for 1 K+.[4] By taking up glutamate and GABA, astrocytes can synthesize glutamine for neurons to use as precursors for more glutamate and glutathione.[4] In doing so, the astrocytes not only cycle ammonia through the nervous system and prevent toxic overload but also prevent excitotoxicity by ridding the extracellular environment of excitatory neurotransmitters that, if left alone, would induce pathological action potentials to fire in neurons, ultimately killing them in the process.[4][5]
Astrocytes also specialize in neuroprotection, neural repair mechanisms, and axonal growth promotion. Although astrocytes already defend the CNS by way of metabolic and homeostatic regulation, they also satisfy an immune function in the brain. Astrocytes can release and respond to both pro- and anti-inflammatory signals, such as cytokines and chemokines. Specifically, astrocytes release substances such as interleukin-6 (IL-6) and tissue necrosis factors (TNF) alpha and beta to promote immune responses in the brain. Astrocytes can also respond to inflammatory signals by undergoing a process known as astrogliosis.[5][6] In astrogliosis, astrocytes become reactive and form fibrous scars that quarantine injured tissues, mitigating the spread of damage.[5][6] Aside from preventing the incursion and spread of neural damage, astrocytes facilitate regeneration and repair of these damaged tissues. Though previous studies claimed that astrocyte scars impeded the regrowth of damaged axons, newer research has shown that astrocytes release axon growth-promoting factors such as nerve growth factor, brain-derived trophic factor, NT3, BDNF, and chondroitin sulfate proteoglycans 4 and 5 (CSPG-4 and CSPG-5) that are essential for axon growth both before and after injury.[6][7] Also, astrocytes promote the formation and maturation of synapses by releasing factors such as thrombospondins (TSPs) 1 and 2 as well as glypican-4 and 6, respectively.[5]
Tissue Preparation
To view astrocytes, histologists have traditionally stained neural tissue samples with hematoxylin and eosin (H&E) or Holzer stains. Newer stains are now available for neural tissue preparation and visualization of astrocytes.
Histochemistry and Cytochemistry
Glial fibrillary acidic protein (GFAP) is the most popular histochemical and cytochemical marker used to identify astrocytes in neural tissue samples.[1][2][8] GFAP expression is not restricted to just astrocytes; other cells, such as proliferating stem cells, express GFAP in the CNS but are morphologically dissimilar to astrocytes.[2][8] To account for this lack of specificity, investigators have identified other markers that can pinpoint astrocytes in neuroimaging studies. These markers include the gap junction protein connexin 43 (Cx43), the cytoplasmic protein S100 beta, the aldehyde dehydrogenase Aldh1L1, and the glutamate transporters GLAST/EAAT-1 and GLT-1/EAAT-2.[2][5][8] While morphology remains the only reliable means of differentiating between astrocyte types,[2][8] markers such as connexin 30 (Cx30), GLAST/EAAT-1, GLT-1/EAAT-2, and potassium channel Kir4, can help identify certain astrocyte types since they preferentially localize around gray matter, white matter, or specific brain regions.[8]
Microscopy, Light
When viewed under a light microscope, astrocytes appear as large nuclei with minimal surrounding cytoplasm, and some sharp protrusions representing primary foot processes whose radial arrangement around the large, circular nucleus gives astrocytes the star-like appearance from whence they derive their name. Holzer staining shows the foot processes better than H&E staining. The best results for light microscopy, however, come from using fluorescent immunocytochemical markers such as GFAP, GLAST/EAAT1, and GLT-1/EAAT2.[8]
In modern practice, conventional light microscopy has fallen out of favor for viewing astrocytes because finer astrocytic foot processes are not readily distinguishable from other fine branching structures such as dendrites or extracellular matrix.[9] These finer foot processes, known as peripheral astrocyte processes (PAPs), constitute the majority of an astrocyte’s volume. Obtaining a clear microscopic view of them is essential for identifying certain astrocyte pathologies.[9] Light microscopes also lack sufficient image definition to determine the type of astrocyte being viewed.[8][9][10] These drawbacks reduce the practicality of conventional light microscopy in astrocyte studies. Special light microscopy techniques are still widely used, however (e.g., immunofluorescent light microscopy to image astrocytes and time-lapse light microscopy to track movements in astroglial protrusions).[8][10]
Microscopy, Electron
With its higher-resolution images, electron microscopy has become the preferred method for viewing astrocytes and their finer processes.[8][10] Under an electron microscope, astrocytes appear as heterogeneous, dense cell bodies with round or oval nuclei, glycogen granules, and many radially-projecting filaments with numerous leaflets.[11][12] Pathological astrocytes change their appearance depending on how much time has elapsed since the initial injury. Acutely, reactive astrocytes synthesize more glycogen granules and mitochondria, become more fibrous as they form more intermediate filaments in their cytoplasm, and have more compacted genetic material in their nuclei.[12] Once scars form, reactive astrocytes lose their extra glycogen granules and mitochondria but become dense with cytoplasmic intermediate filaments.[12] Immunoelectron microscopy of astrocytes is also possible, meaning light microscopes are not required when viewing astrocyte samples with immunostaining.[9]
The current limitations of electron microscopy for viewing astrocytes stem from the fact that electron microscopy requires a large time investment and is unusable on live tissue.[10] Although it is viable in conjunction with other imaging techniques to detect and monitor structural changes in the brain after long-term potentiation and other changes post-stimulation, electron microscopy itself cannot track these changes as they occur.[10] For these reasons, electron microscopy has not fully replaced confocal and light microscopy with immunostaining for in vivo astrocyte imaging.[9][10][13]
Pathophysiology
Astrocyte pathologies tend to manifest in two main forms: astrocytopathy and astrogliosis.[5] Astrocytopathy includes any genetic, traumatic, toxic, or other insult that can alter the existing functions or capabilities of astrocytes.[5] Such pathologies affect the astrocyte network's ability to carry out one or more of its many essential functions, which can lead to a variety of neurological diseases.
The category of astrogliosis is understood to be a neuroimmunological malfunction. Reactive astrogliosis is a complex process in which signals from microglia, synapses, neurons, and sites of injury cause astrocytes to undergo rapid remodeling.[5][6] Astrocytes can react in this way to a variety of ions, transcription factors, metabolic molecules, and even pro-inflammatory signals such as interleukin-6 (IL-6), transforming growth factor-alpha (TGF-alpha), leukemia inhibitory factor (LIF), and ciliary neurotrophic factor (CNTF).[5] These signals promote increased expression of GFAP and vimentin, hypertrophy of foot processes, and expression of pro-inflammatory gene products that allow astrocytes to perform neuroprotective duties.[5] Studies have shown that astrogliosis tends to isolate damaged CNS tissues while protecting healthy tissue, creating glial scars, and repairing damage to the blood-brain barrier while guiding post-traumatic neuroplasticity.[5] The exact response depends on the specific brain region in which astrogliosis gets triggered, but as with many immune responses, there is a risk for pathological damage if the process becomes dysregulated.[5]
Clinical Significance
Because astrocytes fulfill many essential functions, their dysfunction has implicated them in several neurological disorders. Examples include Alzheimer disease (AD),[5][14] epilepsy, Huntington disease, Parkinson disease, multiple sclerosis, amyotrophic lateral sclerosis (ALS), and Alexander disease. Astrocytes also become more reactive during the normal aging process.
In Alzheimer disease, increasing cleavage of amyloid precursor protein (APP) by beta and gamma secretases (instead of alpha-secretase, which normally cleaves APP into a nontoxic form that the body can clear) cause abnormal beta-amyloid plaques to form.[14] In the healthy brain, normal astrocytes become reactive when exposed to amyloid plaques, triggering the astrocytes to release proteases and other substances through their foot processes whose function is to degrade and clear amyloid plaques from the CNS.[5][14] Alzheimer disease, however, is characterized by the pathological accumulation of amyloid plaques accumulate that cause astrocytes to begin deteriorating, particularly in the locus coeruleus, a main adrenergic center of the brain where astrocytes carry out the essential functions of glycogen metabolism, synapse maintenance, and calcium signaling.[5][14] Abnormal astrocytes also express high volumes of APP and apolipoprotein E (Apo-E), both of which are hallmarks and risk factors for Alzheimer disease.[5] Thus, even though the brain needs astrocytes to remove amyloid plaques, too high a concentration of astrocytes interferes with their adrenergic signaling, adversely affecting homeostasis and signaling support, which ultimately worsens the cognitive deficits observed in Alzheimer dementia.[5][14]
Astrocytes affect epilepsy and Huntington disease by regulating homeostatic conditions in the CNS. Mutations in the Kir4.1, aquaporin-4 (AQP4), GLT-1, or GLAST genes contribute to epilepsy by impairing astrocytes’ ability to regulate potassium, water, and glutamate levels in the extracellular fluid within the CNS.[5][15] Specifically, inactivating mutations in Kir4.1 and AQP4 - or down-regulation of their expression - limits uptake and buffering of potassium in extracellular fluid resulting in potassium overload that causes positive shifts in local membrane potential, allows uncontrolled ion conduction, and ultimately triggers uncontrolled action potentials that culminate in clinical seizures.[4][5][15] In the case of GLT-1 and GLAST, loss-of-function mutations or down-regulation derail normal uptake and recycling of glutamate, creating an excitotoxic environment for neurons that generates uncontrolled action potentials and seizures.[4][5][15] A similar pathological process occurs in Huntington disease: impaired Kir4.1 and EAAT2 transporters - coupled with rampant glutamate production from increased pyruvate carboxylase expression - lead to excitotoxic extracellular environments that kill neurons in the basal ganglia, leading to the characteristic motor and cognitive decline seen in Huntington syndrome.[5]
Astrocytes also initiate neuronal death in parkinsonism and can trigger demyelinating disease. In idiopathic Parkinson disease, neurons die in the pars compacta of the substantia nigra when alpha-synuclein deposits accumulate. These alpha-synuclein deposits form a histological marker useful in diagnosing parkinsonism.[14] The astrocytic variant of monoamine oxidase B converts 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) into its neurotoxic form, 1-methyl-4-phenylpyridium (MPP), which can trigger the neurodegenerative processes seen in parkinsonism.[4][14] Because alpha-synuclein collects in reactive astrocytes in the substantia nigra pars compacta, researchers have suggested that these stressed astrocytes may trigger the immune system to destroy surrounding neurons by releasing pro-inflammatory signals.[14] An atypical form of parkinsonism (corticobasal degeneration) gets triggered by abnormal accumulation of tau protein (4-repeat tau isoforms) within the white matter in the cerebral hemispheres and subcortical nuclei. Corticobasal degeneration also affects glial cells, causing astrocytic plaques formed by tau-immunoreactive astrocytic processes.[16] Astrocytes' ability to trigger inflammatory responses in the CNS could play a similar role in multiple sclerosis by attracting CD4+ T-lymphocytes and signaling them with interleukins to attack nearby myelin sheaths.[14] Multiple sclerosis has indeed become an exciting area for emerging research on astrocytes' critical importance to CNS function. Once thought to be bystanders in the multiple sclerosis disease process, astrocytes have now been shown to drive the formation of demyelinating lesions by recruiting inflammatory cells that cross the blood-brain barrier.[17][18] In another demyelinating disease that is much less common than multiple sclerosis (neuromyelitis optica spectrum disorder, or NMOSD), the aquaporin-4 water channel is expressed on astrocytic foot processes in areas of the brain that surround the ventricles and lack a blood-brain barrier.[19] One of these areas, known as the area postrema, when affected by NMOSD, can lead to intractable nausea, vomiting, and hiccups.
Alexander disease and inherited forms of amyotrophic lateral sclerosis (ALS) are examples of diseases that stem from errors in astrocyte genetics. Alexander disease is a genetic astrogliopathy in which glial fibrillary acidic protein (GFAP) mutations disrupt astrocytes' formation of intermediate filaments, causing symmetrical white matter lesions in the frontal lobe and sometimes elsewhere in the brain.[5] These mutations form GFAP aggregates known as Rosenthal fibers, which are composed of vimentin, aB-crystallin, and Hsp27.[5] Inherited forms of amyotrophic lateral sclerosis represent additional astrocyte-dependent genetic diseases. Inherited mutations in SOD1, including the G93A mutation, disturb the normal function of both astrocytes and neurons.[14][20] When mutated SOD1 aggregates accumulate, activated astrocytes begin to release pro-inflammatory cytokines (e.g., IL-6 and TGF-beta) that impair communication between neurons and astrocytes.[14] Mechanisms include faulty lactate transport needed to meet increasing neuronal metabolic demands, loss of GLT-1 glutamate transporters, changes in AMPA receptor subunit GluR2 expression, and autoimmune targeting of neurons by reactive astrocytes and microglia via STAT3 signaling errors.[14] The result is a neurotoxic environment which leads to the inexorable motor neuron impairment seen in ALS.[4][14]
As the traditional "neuron-centric" view of the brain gives way to a more comprehensive understanding that includes the complex contributions of astrocytes and other non-neuronal cells, future research is turning to the astrocyte's role in new therapeutic approaches. As investigators learn more about how astrocytes function malfunction, new insights may enable treatment strategies that would not have been possible in decades past.[3] Exploring astrocytes' role in the SOD1 gene mutations responsible for heritable forms of ALS,[20][21] for instance, may suggest novel therapies to treat this neurologically devastating disease. Future research will likely continue to target astrocytes' potential roles in protecting the CNS from damage - as well as promoting regeneration of existing brain lesions - caused by a wide variety of neurological disease states.
Disclaimer: This scholarly activity was supported (in whole or in part) by HCA and/or an HCA-affiliated entity. The views expressed in this publication represent those of the author(s) and do not necessarily represent the official views of HCA or any of its affiliated entities.