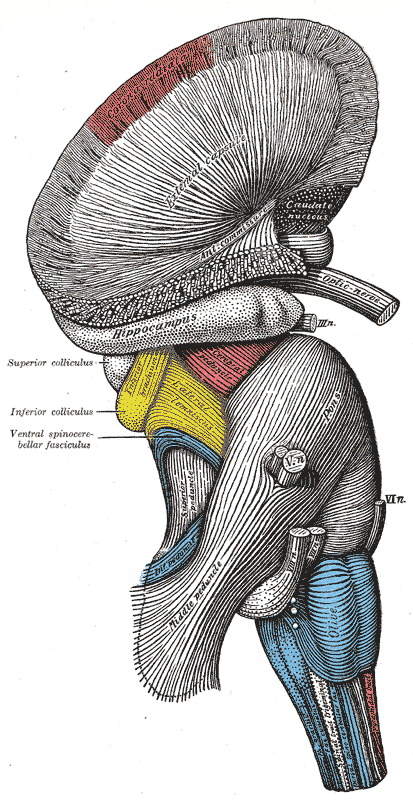
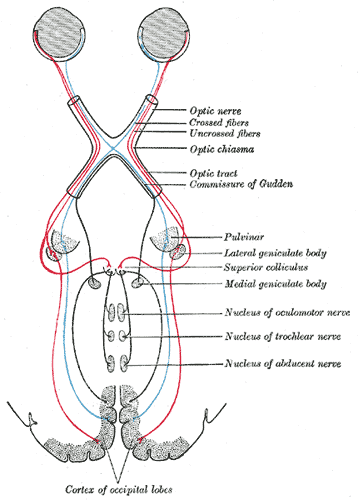
[1]
Lane RH, Allman JM, Kaas JH, Miezin FM. The visuotopic organization of the superior colliculus of the owl monkey (Aotus trivirgatus) and the bush baby (Galago senegalensis). Brain research. 1973 Oct 12:60(2):335-49
[PubMed PMID: 4202853]
[2]
Gandhi NJ, Katnani HA. Motor functions of the superior colliculus. Annual review of neuroscience. 2011:34():205-31. doi: 10.1146/annurev-neuro-061010-113728. Epub
[PubMed PMID: 21456962]
[4]
Sparks DL, Gandhi NJ. Single cell signals: an oculomotor perspective. Progress in brain research. 2003:142():35-53
[PubMed PMID: 12693253]
Level 3 (low-level) evidence
[5]
Thieu MK, Ayzenberg V, Lourenco SF, Kragel PA. Visual looming is a primitive for human emotion. bioRxiv : the preprint server for biology. 2024 Jan 3:():. pii: 2023.08.29.555380. doi: 10.1101/2023.08.29.555380. Epub 2024 Jan 3
[PubMed PMID: 37693448]
[6]
Prabhu NG, Himmelbach M. Activity in the human superior colliculus associated with reaching for tactile targets. NeuroImage. 2023 Oct 15:280():120322. doi: 10.1016/j.neuroimage.2023.120322. Epub 2023 Aug 15
[PubMed PMID: 37586443]
[7]
Yu G, Katz LN, Quaia C, Messinger A, Krauzlis RJ. Short-latency preference for faces in the primate superior colliculus. bioRxiv : the preprint server for biology. 2023 Sep 9:():. pii: 2023.09.06.556401. doi: 10.1101/2023.09.06.556401. Epub 2023 Sep 9
[PubMed PMID: 37886488]
[8]
Thompson J, Lovicu F, Ziman M. The role of Pax7 in determining the cytoarchitecture of the superior colliculus. Development, growth & differentiation. 2004 Jun:46(3):213-8
[PubMed PMID: 15206952]
[9]
Omi M, Nakamura H. Engrailed and tectum development. Development, growth & differentiation. 2015 Feb:57(2):135-45. doi: 10.1111/dgd.12197. Epub 2015 Feb 25
[PubMed PMID: 25716935]
[10]
Tatu L, Moulin T, Bogousslavsky J, Duvernoy H. Arterial territories of human brain: brainstem and cerebellum. Neurology. 1996 Nov:47(5):1125-35
[PubMed PMID: 8909417]
[11]
Ruchalski K, Hathout GM. A medley of midbrain maladies: a brief review of midbrain anatomy and syndromology for radiologists. Radiology research and practice. 2012:2012():258524. doi: 10.1155/2012/258524. Epub 2012 May 22
[PubMed PMID: 22693668]
[12]
van Lessen M, Shibata-Germanos S, van Impel A, Hawkins TA, Rihel J, Schulte-Merker S. Intracellular uptake of macromolecules by brain lymphatic endothelial cells during zebrafish embryonic development. eLife. 2017 May 12:6():. pii: e25932. doi: 10.7554/eLife.25932. Epub 2017 May 12
[PubMed PMID: 28498105]
[13]
Edwards SB, Henkel CK. Superior colliculus connections with the extraocular motor nuclei in the cat. The Journal of comparative neurology. 1978 May 15:179(2):451-67
[PubMed PMID: 641226]
Level 2 (mid-level) evidence
[14]
Gaymard B, François C, Ploner CJ, Condy C, Rivaud-Péchoux S. A direct prefrontotectal tract against distractibility in the human brain. Annals of neurology. 2003 Apr:53(4):542-5
[PubMed PMID: 12666125]
[15]
Liu APY, Harreld JH, Jacola LM, Gero M, Acharya S, Ghazwani Y, Wu S, Li X, Klimo P Jr, Gajjar A, Chiang J, Qaddoumi I. Tectal glioma as a distinct diagnostic entity: a comprehensive clinical, imaging, histologic and molecular analysis. Acta neuropathologica communications. 2018 Sep 25:6(1):101. doi: 10.1186/s40478-018-0602-5. Epub 2018 Sep 25
[PubMed PMID: 30253793]
[16]
Lapras C, Bognar L, Turjman F, Villanyi E, Mottolese C, Fischer C, Jouvet A, Guyotat J. Tectal plate gliomas. Part I: Microsurgery of the tectal plate gliomas. Acta neurochirurgica. 1994:126(2-4):76-83
[PubMed PMID: 8042559]
[17]
Kurtz D, Butter CM. Impairments in visual discrimination performance and gaze shifts in monkeys with superior colliculus lesions. Brain research. 1980 Aug 25:196(1):109-24
[PubMed PMID: 6772274]
[18]
MacKinnon DA, Gross CG, Bender DB. A visual deficit after superior colliculus lesions in monkeys. Acta neurobiologiae experimentalis. 1976:36(1-2):169-80
[PubMed PMID: 823800]
[19]
Goodale MA, Murison RC. The effects of lesions of the superior colliculus on locomotor orientation and the orienting reflex in the rat. Brain research. 1975 May 2:88(2):243-61
[PubMed PMID: 1148825]