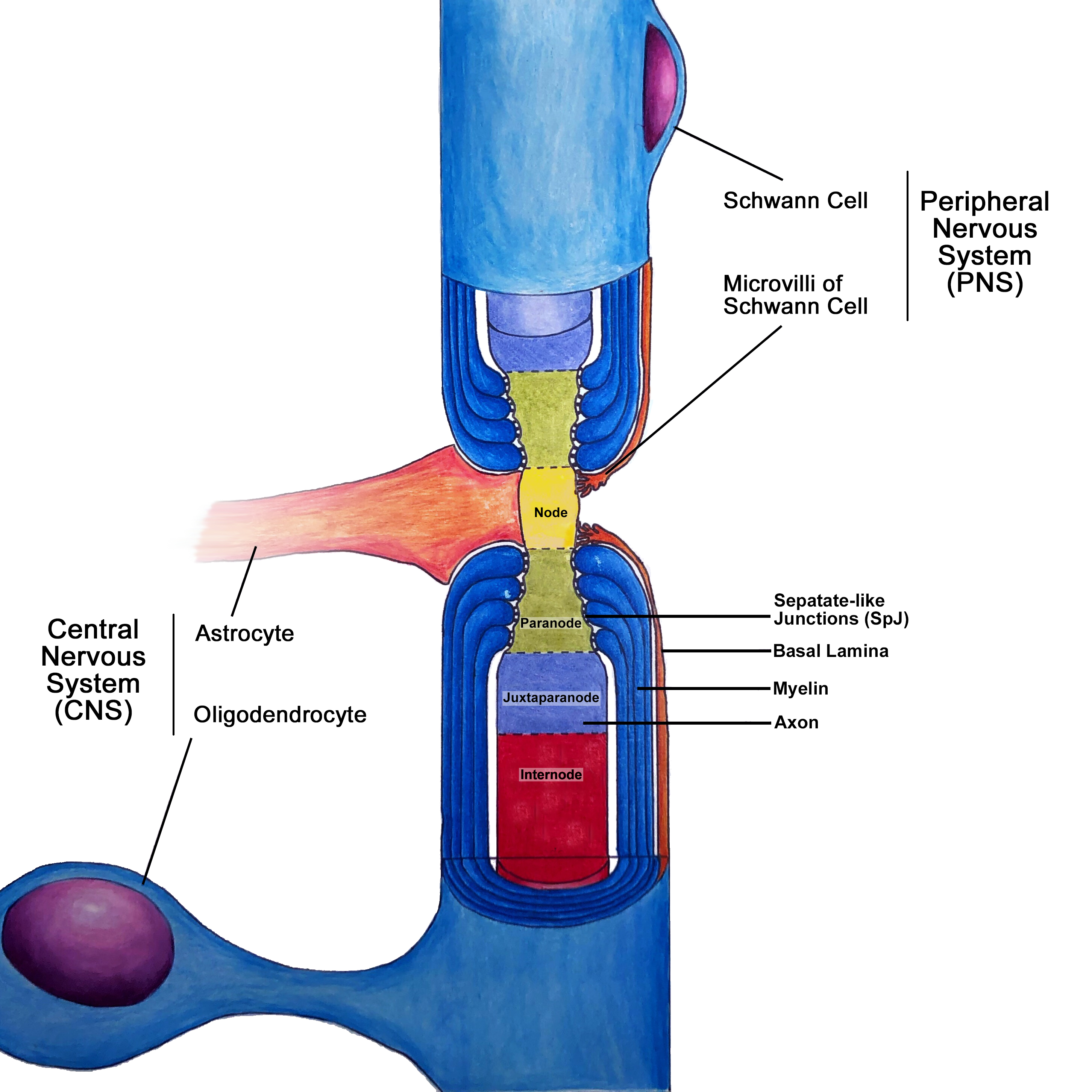
[1]
Arancibia-Carcamo IL, Attwell D. The node of Ranvier in CNS pathology. Acta neuropathologica. 2014 Aug:128(2):161-75. doi: 10.1007/s00401-014-1305-z. Epub 2014 Jun 10
[PubMed PMID: 24913350]
[2]
Davis JQ, Lambert S, Bennett V. Molecular composition of the node of Ranvier: identification of ankyrin-binding cell adhesion molecules neurofascin (mucin+/third FNIII domain-) and NrCAM at nodal axon segments. The Journal of cell biology. 1996 Dec:135(5):1355-67
[PubMed PMID: 8947556]
[3]
Poliak S, Peles E. The local differentiation of myelinated axons at nodes of Ranvier. Nature reviews. Neuroscience. 2003 Dec:4(12):968-80
[PubMed PMID: 14682359]
[4]
Rasband MN, Peles E. The Nodes of Ranvier: Molecular Assembly and Maintenance. Cold Spring Harbor perspectives in biology. 2015 Sep 9:8(3):a020495. doi: 10.1101/cshperspect.a020495. Epub 2015 Sep 9
[PubMed PMID: 26354894]
Level 3 (low-level) evidence
[6]
Zhang YP, Huang QL, Zhao CM, Tang JL, Wang YL. GM1 improves neurofascin155 association with lipid rafts and prevents rat brain myelin injury after hypoxia-ischemia. Brazilian journal of medical and biological research = Revista brasileira de pesquisas medicas e biologicas. 2011 Jun:44(6):553-61
[PubMed PMID: 21670940]
[7]
Snaidero N, Simons M. Myelination at a glance. Journal of cell science. 2014 Jul 15:127(Pt 14):2999-3004. doi: 10.1242/jcs.151043. Epub
[PubMed PMID: 25024457]
[8]
Fex Svennigsen A, Dahlin LB. Repair of the Peripheral Nerve-Remyelination that Works. Brain sciences. 2013 Aug 2:3(3):1182-97. doi: 10.3390/brainsci3031182. Epub 2013 Aug 2
[PubMed PMID: 24961524]
[9]
M F G, M M, S H, Khan WS. Peripheral nerve injury: principles for repair and regeneration. The open orthopaedics journal. 2014:8():199-203. doi: 10.2174/1874325001408010199. Epub 2014 Jun 27
[PubMed PMID: 25067975]
[10]
Howell OW, Palser A, Polito A, Melrose S, Zonta B, Scheiermann C, Vora AJ, Brophy PJ, Reynolds R. Disruption of neurofascin localization reveals early changes preceding demyelination and remyelination in multiple sclerosis. Brain : a journal of neurology. 2006 Dec:129(Pt 12):3173-85
[PubMed PMID: 17041241]
[11]
Griggs RB, Yermakov LM, Susuki K. Formation and disruption of functional domains in myelinated CNS axons. Neuroscience research. 2017 Mar:116():77-87. doi: 10.1016/j.neures.2016.09.010. Epub 2016 Oct 4
[PubMed PMID: 27717670]
[12]
Traboulsee AL, Li DK. The role of MRI in the diagnosis of multiple sclerosis. Advances in neurology. 2006:98():125-46
[PubMed PMID: 16400831]
Level 3 (low-level) evidence
[13]
Kieseier BC. The mechanism of action of interferon-β in relapsing multiple sclerosis. CNS drugs. 2011 Jun 1:25(6):491-502. doi: 10.2165/11591110-000000000-00000. Epub
[PubMed PMID: 21649449]
[14]
Kira JI, Yamasaki R, Ogata H. Anti-neurofascin autoantibody and demyelination. Neurochemistry international. 2019 Nov:130():104360. doi: 10.1016/j.neuint.2018.12.011. Epub 2018 Dec 22
[PubMed PMID: 30582947]
[15]
van den Berg B, Walgaard C, Drenthen J, Fokke C, Jacobs BC, van Doorn PA. Guillain-Barré syndrome: pathogenesis, diagnosis, treatment and prognosis. Nature reviews. Neurology. 2014 Aug:10(8):469-82. doi: 10.1038/nrneurol.2014.121. Epub 2014 Jul 15
[PubMed PMID: 25023340]
[16]
Hiraga A, Mori M, Ogawara K, Kojima S, Kanesaka T, Misawa S, Hattori T, Kuwabara S. Recovery patterns and long term prognosis for axonal Guillain-Barré syndrome. Journal of neurology, neurosurgery, and psychiatry. 2005 May:76(5):719-22
[PubMed PMID: 15834034]
[17]
Kieseier BC, Mathey EK, Sommer C, Hartung HP. Immune-mediated neuropathies. Nature reviews. Disease primers. 2018 Oct 11:4(1):31. doi: 10.1038/s41572-018-0027-2. Epub 2018 Oct 11
[PubMed PMID: 30310069]
[18]
Mel'nikov RA, Goshchitskiĭ LG, Kovalev VK. [Clinical manifestations of squamous cell carcinoma of the rectum]. Voprosy onkologii. 1984:30(8):76-83
[PubMed PMID: 6485288]
[19]
Palve SS, Palve SB. Impact of Aging on Nerve Conduction Velocities and Late Responses in Healthy Individuals. Journal of neurosciences in rural practice. 2018 Jan-Mar:9(1):112-116. doi: 10.4103/jnrp.jnrp_323_17. Epub
[PubMed PMID: 29456354]