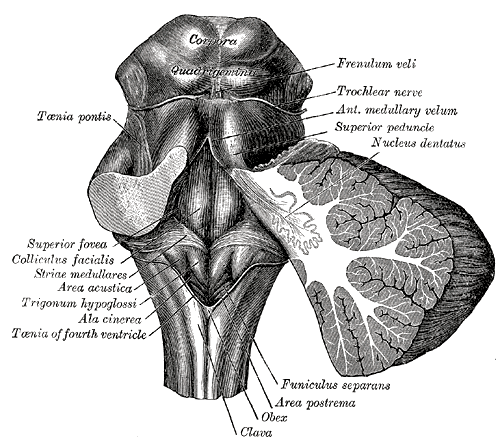
[1]
Sarnat HB, Flores-Sarnat L, Boltshauser E. Area Postrema: Fetal Maturation, Tumors, Vomiting Center, Growth, Role in Neuromyelitis Optica. Pediatric neurology. 2019 May:94():21-31. doi: 10.1016/j.pediatrneurol.2018.12.006. Epub 2018 Dec 25
[PubMed PMID: 30797593]
[2]
Beddok A, Faivre JC, Coutte A, Guévelou JL, Welmant J, Clavier JB, Guihard S, Janoray G, Calugaru V, Pointreau Y, Lacout A, Salleron J, Lefranc M, Hasboun D, Duvernoy HM, Thariat J. Practical contouring guidelines with an MR-based atlas of brainstem structures involved in radiation-induced nausea and vomiting. Radiotherapy and oncology : journal of the European Society for Therapeutic Radiology and Oncology. 2019 Jan:130():113-120. doi: 10.1016/j.radonc.2018.08.003. Epub 2018 Aug 29
[PubMed PMID: 30172454]
[3]
Miller AD, Leslie RA. The area postrema and vomiting. Frontiers in neuroendocrinology. 1994 Dec:15(4):301-20
[PubMed PMID: 7895890]
[4]
Gokozan HN, Baig F, Corcoran S, Catacutan FP, Gygli PE, Takakura AC, Moreira TS, Czeisler C, Otero JJ. Area postrema undergoes dynamic postnatal changes in mice and humans. The Journal of comparative neurology. 2016 Apr 15:524(6):1259-69. doi: 10.1002/cne.23903. Epub 2015 Dec 17
[PubMed PMID: 26400711]
Level 2 (mid-level) evidence
[5]
Pangestiningsih TW, Hendrickson A, Sigit K, Sajuthi D, Nurhidayat, Bowden DM. Development of the area postrema: an immunohistochemical study in the macaque. Brain research. 2009 Jul 14:1280():23-32. doi: 10.1016/j.brainres.2009.05.028. Epub 2009 May 19
[PubMed PMID: 19460361]
[6]
Duvernoy H, Koritké JG, Monnier G, Jacquet G. [Vascularization of the area postrema and of the dorsal side of the medulla oblongata in the human]. Zeitschrift fur Anatomie und Entwicklungsgeschichte. 1972:138(1):41-66
[PubMed PMID: 4638677]
[7]
Weller RO, Djuanda E, Yow HY, Carare RO. Lymphatic drainage of the brain and the pathophysiology of neurological disease. Acta neuropathologica. 2009 Jan:117(1):1-14. doi: 10.1007/s00401-008-0457-0. Epub 2008 Nov 11
[PubMed PMID: 19002474]
[8]
Maolood N, Meister B. Protein components of the blood-brain barrier (BBB) in the brainstem area postrema-nucleus tractus solitarius region. Journal of chemical neuroanatomy. 2009 May:37(3):182-95. doi: 10.1016/j.jchemneu.2008.12.007. Epub 2008 Dec 25
[PubMed PMID: 19146948]
[9]
Vigier D, Rouvière A. Afferent and efferent connections of the area postrema demonstrated by the horseradish peroxidase method. Archives italiennes de biologie. 1979 Nov:117(4):325-39
[PubMed PMID: 550738]
[10]
Babic T, Browning KN. The role of vagal neurocircuits in the regulation of nausea and vomiting. European journal of pharmacology. 2014 Jan 5:722():38-47. doi: 10.1016/j.ejphar.2013.08.047. Epub 2013 Oct 31
[PubMed PMID: 24184670]
[11]
Longatti P, Porzionato A, Basaldella L, Fiorindi A, De Caro P, Feletti A. The human area postrema: clear-cut silhouette and variations shown in vivo. Journal of neurosurgery. 2015 May:122(5):989-95. doi: 10.3171/2014.11.JNS14482. Epub 2015 Jan 16
[PubMed PMID: 25594320]
[12]
Spencer CM, Eckel LA, Nardos R, Houpt TA. Area postrema lesions attenuate LiCl-induced c-Fos expression correlated with conditioned taste aversion learning. Physiology & behavior. 2012 Jan 18:105(2):151-60. doi: 10.1016/j.physbeh.2011.08.022. Epub 2011 Aug 24
[PubMed PMID: 21889521]
[13]
Travagli RA, Anselmi L. Vagal neurocircuitry and its influence on gastric motility. Nature reviews. Gastroenterology & hepatology. 2016 Jul:13(7):389-401. doi: 10.1038/nrgastro.2016.76. Epub 2016 May 25
[PubMed PMID: 27251213]
[14]
Camara-Lemarroy CR, Burton JM. Area postrema syndrome: A short history of a pearl in demyelinating diseases. Multiple sclerosis (Houndmills, Basingstoke, England). 2019 Mar:25(3):325-329. doi: 10.1177/1352458518813105. Epub 2018 Nov 22
[PubMed PMID: 30463481]