Introduction
The dorsal spinocerebellar tract (DSCT), also known as the posterior spinocerebellar tract or Flechsig tract, is a somatosensory part of the sensory nervous system that relays unconscious proprioceptive information from the lower limbs and trunk of the body to the cerebellum. Sensory information is recognized as an electrical signal and is transmitted via various tracts throughout the nervous system. Proprioception is the ability to sense position and perform movements of different parts of the body. Proprioceptors, a type of mechanoreceptor, are responsible for recognizing proprioceptive signals. Golgi muscle tendons and muscle spindles are two main types of proprioceptors that detect muscle strain and muscle contraction, respectively.[1]
Structure and Function
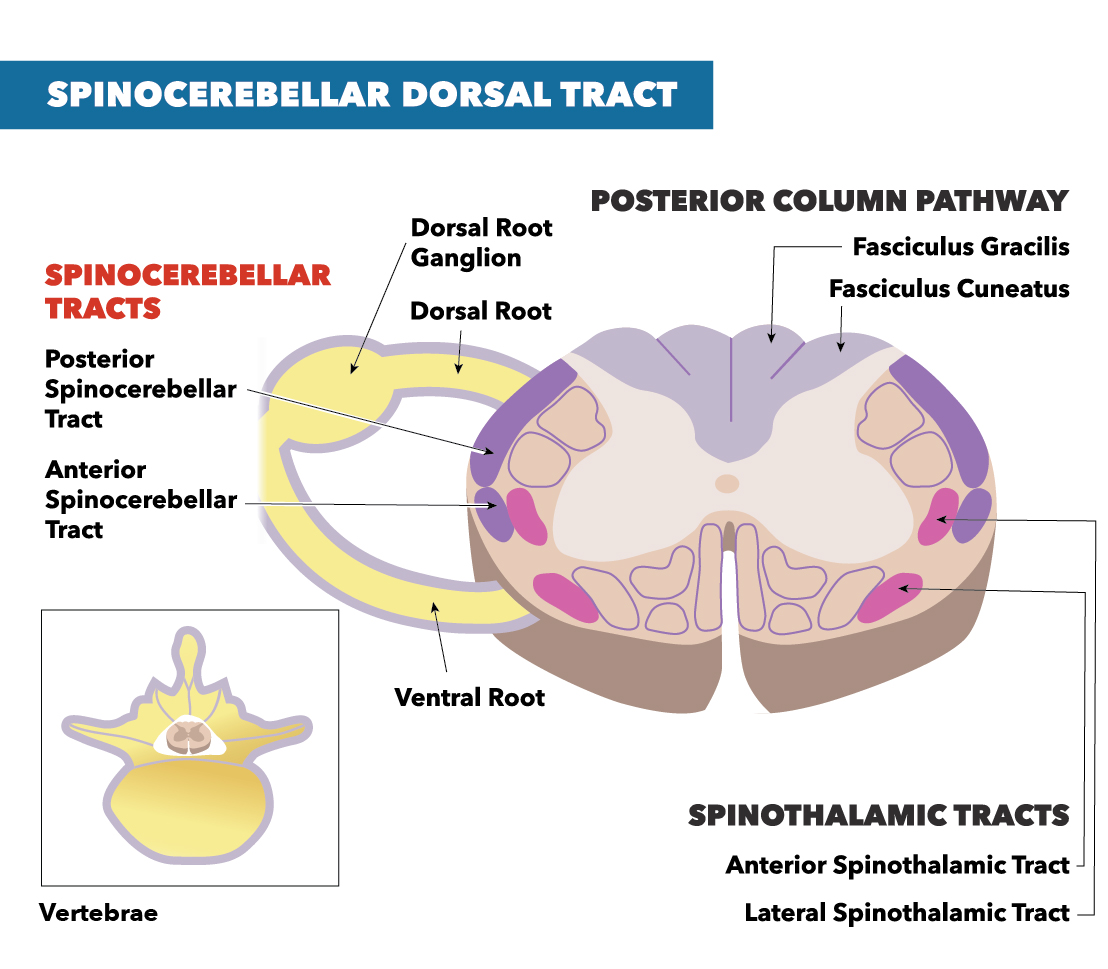
The DSCT transmits proprioception information from Golgi tendon organs and muscle spindles of the trunk and lower limb. Information is relayed from the ipsilateral lower limb and trunk. Each muscle spindle consists of a primary ending and a secondary ending in the middle of the spindle. While the primary endings respond to both velocity and muscle length, the secondary endings can only respond to muscle length. All of the sensory information from the periphery is transmitted to the pseudounipolar neurons in the dorsal root ganglion. The first-order neuron for the dorsal spinocerebellar tract transmits impulses to a specific location in the posterior gray horn known as Clarke’s nucleus. This set of nuclei range from the C8-L3 portions of the spinal cord, collectively referred to as Clarke’s column. The DSCT ascends on the ipsilateral side, enters the cerebellum via the inferior peduncle, and terminates as mossy fibers in the cerebellar cortex. Specifically, the information transmits to the vermis and paravermis in the anterior lobe of the cerebellum. The cerebellum then uses the proprioceptive information to construct and coordinate voluntary movement by knowing the position sense of the lower limbs and the trunk.[2]
The DSCT has a tract that runs parallel with it: the ventral spinocerebellar tract (VSCT). Instead of relaying proprioceptive information, the VSCT relays motor information. Even though both VSCT and DSCT relay information ipsilaterally, the DSCT stays ipsilateral throughout the entire tract, while the VSCT decussates twice. The VSCT decussates first at the level of Clarke’s nucleus and rises contralaterally from the input to the brain. The tract enters through the contralateral superior cerebellar peduncle, where it will decussate back to the ipsilateral side of the input within the cerebellum.[3]
Embryology
Cerebellar-projecting neurons transmit proprioceptive information through the central nervous system. ATOH-1 is a transcription factor implicated in forming these neurons. Research has determined that ATOH-1 lineage neurons contribute to the reception of unconscious proprioceptive information from both the hindbrain (non-Clarke's division of the dorsal spinocerebellar tract) and those from the forelimb (cuneocerebellar tract).[4]
Clinical Significance
The DSCT plays a vital role in the transfer of sensory information of unconscious proprioception. Therefore, damage to DSCT results in ataxia and loss of unconscious proprioception. Ataxia is a condition in which a person lacks the ability to perform voluntary movements due to a loss of muscle control or coordination. Ataxia could result from several conditions ranging from alcohol/drug abuse, medication to genetic and nongenetic neurological diseases. Lower limb conscious and unconscious proprioceptive information is a key component of performing voluntary coordinated movements such as walking. A dysfunctional DSCT leads to an absence of proprioception from the lower limbs, which leads to a loss in coordination, resulting in ataxia.[1]
Anatomically, the spinal cord pathways cluster close to one another. Therefore, when suspecting damage to the DSCT, damage to other tracts nearby should be suspected and tested to confirm function. An isolated injury of the DSCT would be uncommon given the proximity of these critical structures. However, many clinical scenarios can cause DSCT dysfunction: Friedreich ataxia, vitamin E deficiency, tabes dorsalis, Brown-Sequard syndrome, and subacute combined degeneration (vitamin B12 deficiency).
Brown-Sequard syndrome is characterized by damage to one-half (hemisection) of the spinal cord. Consequently, the damage leads to ipsilateral loss of sensation and flaccid paralysis due to damaged lower motor neurons at the level of the lesion. Additionally, the damage leads to ipsilateral loss of proprioception, two-point discrimination, touch, and vibration below the level of the lesion because of the damage to the dorsal column medial lemniscus pathway. The damage to the corticospinal tract also presents with the signs of upper motor neuron damage, such as spastic paralysis below the level of the lesion. The loss of function in the spinothalamic tract causes loss of pain, temperature, and non-discriminative touch on the contralateral side of the body below the level of the lesion.[5]
Friedreich ataxia is an autosomal recessive trinucleotide (GAA) repeat disorder on chromosome 9. The gene affected is an iron-binding protein gene known as the Frataxin gene. This disease is characterized by degeneration of the spinocerebellar tract, dorsal root ganglia, dorsal column, and lateral corticospinal tracts. Additionally, partial loss of mitochondrial function occurs. Patients typically present with ataxia, loss of proprioception, spastic paralysis, and a loss of deep tendon reflexes.[6]
Tabes dorsalis is a condition caused by tertiary syphilis infection due to untreated, prolonged syphilis infection. Tabes dorsalis is characterized by slow demyelination of the dorsal columns and the dorsal roots of the spinal cord. Demyelination causes damage to different tracts, such as the dorsal column medial lemniscus pathway, and the dorsal spinocerebellar tract. Symptoms of tabes dorsalis include diminished reflexes, weakness, ataxia, and Argyll-Robertson pupil.[7]
Subacute combined degeneration is degeneration of the spinocerebellar tracts, corticospinal tracts, and the dorsal column due to a prolonged vitamin B12 deficiency. Physiologically, vitamin B12 is synthesized by microorganisms and stored in the liver on average from three to five years. Because of the extended storage duration, vitamin B12 deficiency develops over long periods. Common causes of vitamin B12 deficiency are malnutrition (veganism, improper diet), lack of intrinsic factor (gastric bypass surgery, pernicious anemia), malabsorption (Crohn's disease), and overgrowth of microorganisms (bacterial, Diphyllobothrium latum). The hematological presentation of vitamin B12 deficiency is macrocytic anemia, similar to the presentation of folate (vitamin B9) deficiency. However, the crucial difference between the two is that neurological symptoms are present in vitamin B12 deficiency.[8]
A vitamin E deficiency could lead to hematological and neurological issues such as hemolytic anemia and demyelination of the dorsal column and the spinocerebellar tract. The neurological symptoms of vitamin E deficiency are similar to the neurological symptoms in vitamin B12 deficiency, but the difference is in the hematological symptoms. There is no evidence of increased serum methylmalonic acid, hypersegmented neutrophils, or megaloblastic anemia in vitamin E deficiency.[9]